
Corynura (Corynura) moscosensis, González-Vaquero, Rocío Ana, Polidori, Carlo & Nieves-Aldrey, José Luis, 2017
|
publication ID |
https://doi.org/ 10.5281/zenodo.246769 |
|
publication LSID |
lsid:zoobank.org:pub:8C86F0AD-B63D-430F-BEA8-4FBE281E7C56 |
|
DOI |
https://doi.org/10.5281/zenodo.6017618 |
|
persistent identifier |
https://treatment.plazi.org/id/EF1787F9-1977-FB50-D9EF-FB51B61FFB50 |
|
treatment provided by |
Plazi |
|
scientific name |
Corynura (Corynura) moscosensis |
| status |
sp. nov. |
Corynura (Corynura) moscosensis n. sp. González-Vaquero
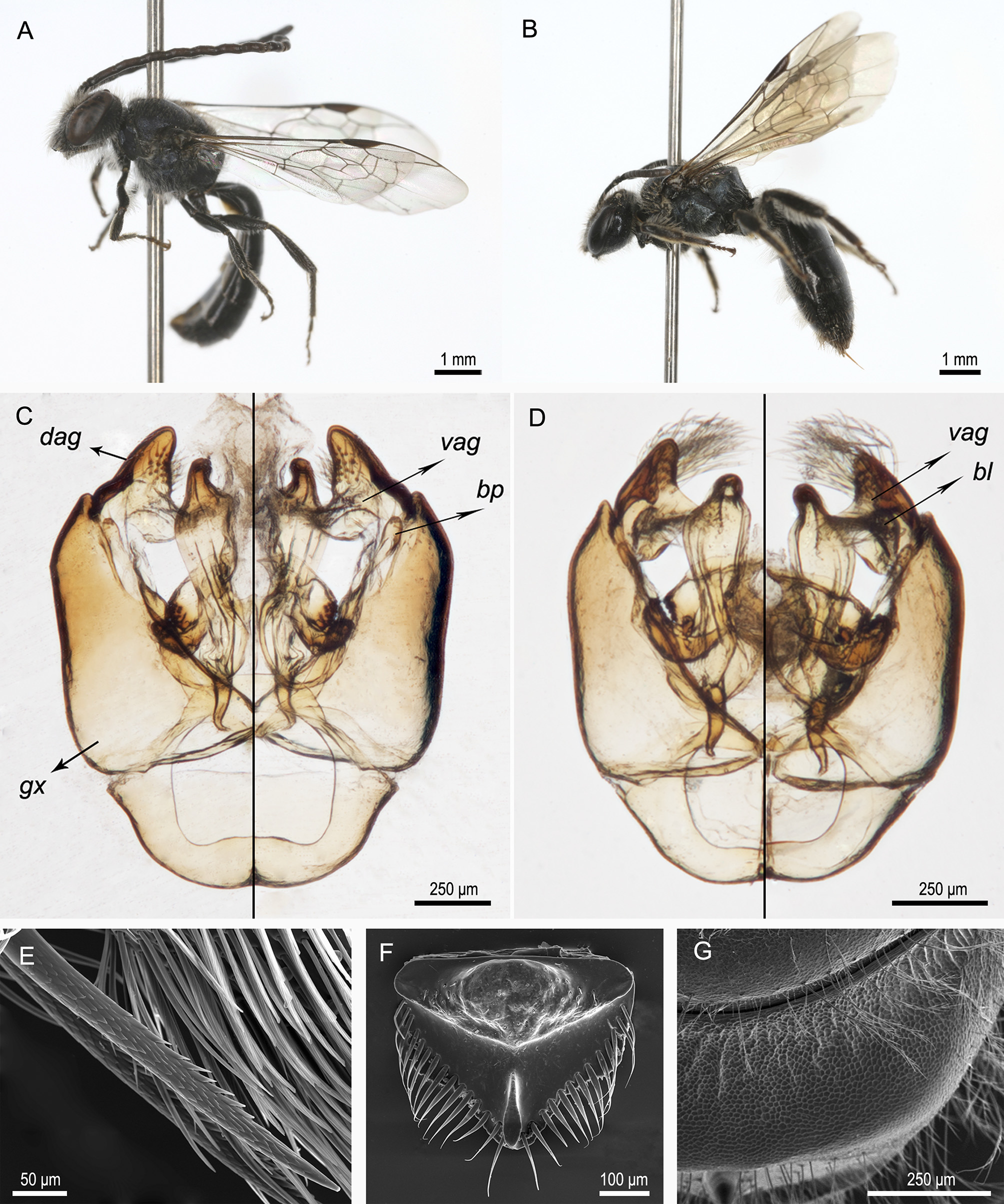
( Figs. 1A View FIGURE 1 –C, 1E–G)
Diagnosis. Corynura moscosensis n. sp. is a moderate-size bee (7.0– 8.4 mm), black with bluish highlights and an entirely dark-brown metasoma. The females of this species can be distinguished from C. chloromelas and other similar Corynura by the following combination of characters: inner hind tibial spur with many short teeth ( Fig. 1 View FIGURE 1 E), malar area at most 1/4 as long as basal mandibular width, apical margin of metapostnotum roughly tessellate, propodeum bearing only plumose hairs, apex of clypeus more weakly tessellate than its base, and S3 bearing mainly simple hairs. Males can be easily identified by the exclusive SP dorsal area almost completely covering the flagellum ( Fig. 2 View FIGURE 2 A), the punctures on disc of mesoscutum separated by more than 3 PD, the posterior border of metapostnotum straight (margin not upraised), T2–T3 with short (0.1 MOD), simple hairs, with a few simple and plumose hairs no longer than 0.4 MOD, and the following characters of the genital capsule: dorsal area of the gonostylus triangular in shape, apical region of the ventral area bearing short and simple setae ( Fig. 1 View FIGURE 1 C: vag).
Description. Male (holotype, Fig. 1A View FIGURE 1 ). Length, 7.7 mm; forewing length, 6.4 mm (paratypes, length, 7.0– 8.4 mm; forewing length, 6.1–6.9 mm).
Color. Black, with bluish highlights on head and mesosoma, except following areas dark brown: apex of mandible, anterior surface of flagellum, legs, tegula, metasoma. Wings hyaline, with veins and pterostigma dark brown, radial vein dark brown to black.
Pubescence. Whitish, with light brown hairs on head and mesoscutum. Head with plumose hairs as long as 2.0 MOD on clypeus, supraclypeal area, upper paraocular area and vertex. Lower part of gena with hairs up to 2.3 MOD. Upper paraocular area, mesoscutum, mesoscutellum and metanotum with dark, short (0.2–0.3 MOD), simple hairs, denser on lower paraocular area. Mesosoma with plumose hairs on mesoscutum and pleura (1.4–1.6 MOD), longer on metanotum (up to 2.2 times MOD). Base of T2–T3 with short (0.1 MOD), simple hairs, scattered with a few simple and plumose hairs as long as 0.4 MOD, longer on T5–T6 (<1.6 MOD); apical margins with short (0.2 MOD) simple hairs. Sterna with simple hairs, denser on S5–S6, as long as 0.4 MOD.
Sculpture: Clypeus with punctures separated by 1.0–2.0 PD, those on supraclypeal area separated by 2.0–4.0 PD. Lower paraocular area with punctation separated by 1.0–1.5 PD. Punctures on disc of mesoscutum separated by 3.0–5.0 PD, denser at sides, those of the mesoscutellum separated by 2.0–3.0 PD. Dorsal surface of metapostnotum with fine radial striae, surpassing midlength of metapostnotum but not reaching apical margin. Terga with punctures separated by 2.0–3.0 PD, sparser (4.0–5.0 PD) towards apex. Surface between punctures equally tessellate throughout body except: tessellation stronger on frons and upper paraocular area, propodeum minutely wrinkled, metasoma substrigulate.
Structure. Head broader than long, width:length = 1.22:1 (paratypes, 1.16–1.24:1). Ratio of lower to upper interocular distance 0.68:1 (paratypes, 0.65–0.69:1). Clypeus broader than long, 1.45:1 (paratypes, 1.42–1.67:1). Ratio of interantennal to antennocular distance, 1.83:1 (paratypes, 1.60–1.84:1). Ratio of posterior interocellar to ocellocular distance, 1.32:1 (paratypes, 1.11–1.32:1). Ratio of length of scape, pedicel, F1 and F2 0.76:0.26:0.34:1 (paratypes, 0.75–0.97:0.21–0.26:0.26–0.34:1). Apical margins of sterna: S1–S4 straight, S5 with a small median notch, S6 produced, with lateral bend. Hidden sterna: S7 long and curved. Posterior margin of S8 with a median, membranous glabrous process. Genital capsule ( Fig. 1 View FIGURE 1 C): Volsella short, apical margin of digitus with a mesal process. Inner dorsal margin of gonocoxite straight, both margins converging towards the gonobase. Dorsal area of gonostylus triangular, more sclerotized than ventral area ( Fig. 1 View FIGURE 1 C: dag). Ventral area of gonostylus mesally directed, with a small, median, sparsely setose process mesally directed, and apical region with tuft of short, simple setae ( Fig. 1 View FIGURE 1 C: vag). Basal process of gonostylus with a few setae ( Fig. 1 View FIGURE 1 C: bp).
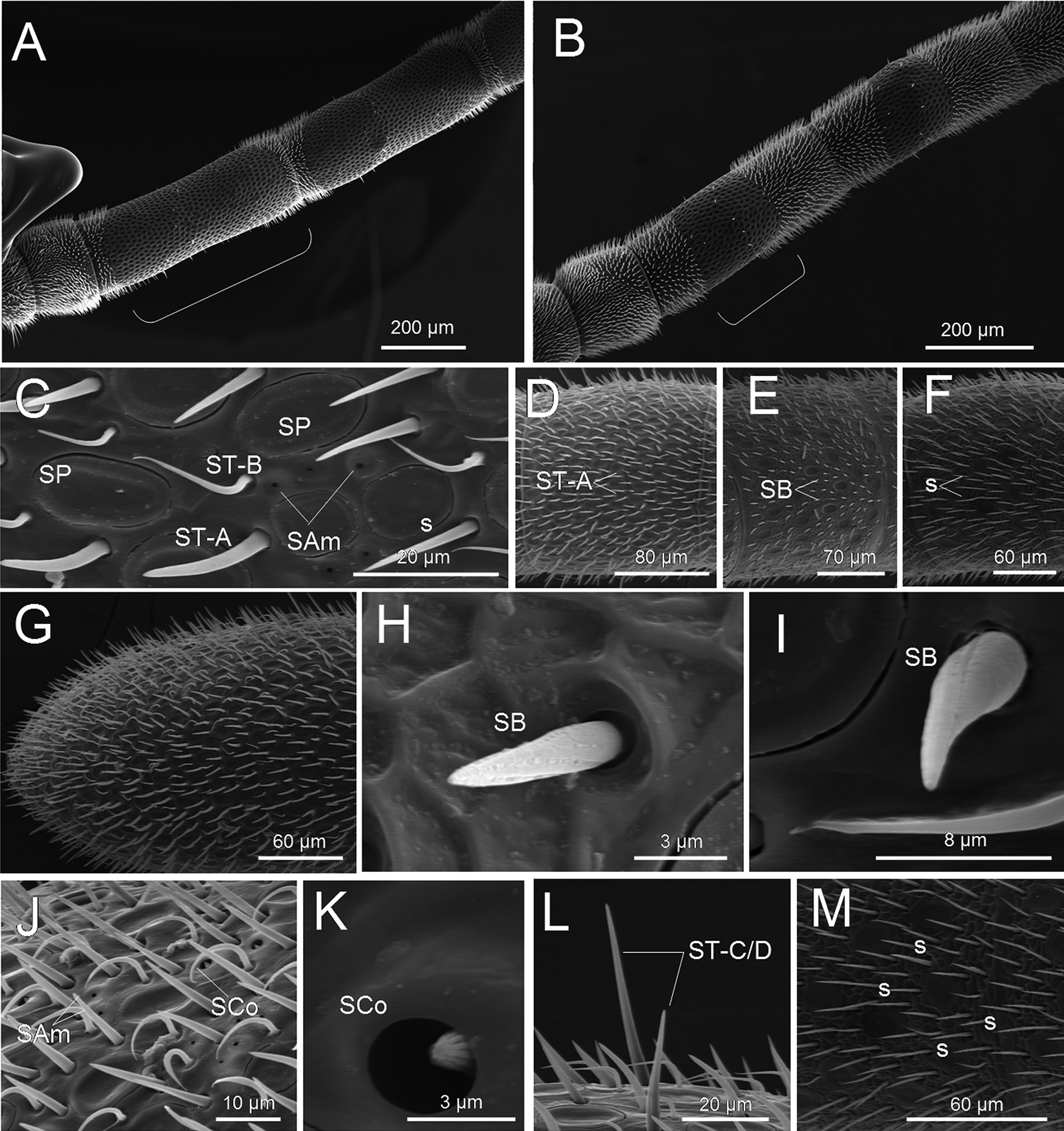
Antennal sensilla. F2–F11 dorsally covered almost exclusively by SP, with a few short ST of one type (ST-C/ D, see below) scattered ( Fig. 2 View FIGURE 2 A). Base and apex of each flagellomere covered by ST of two types (A and B, see below), and a few SP.
Female ( Fig. 1B View FIGURE 1 ). Length, 7.2–7.8 mm; forewing length, 6.1–6.6 mm.
Color. As in male, except mandible with reddish apex.
Pubescence. Light brown, with dark brown hairs on paraocular area. Head with erect, plumose hairs, those on paraocular area and vertex up to 1.7 MOD. Lower part of gena with hairs up to –2.0 MOD. Upper paraocular area with dense, simple, short hairs (0.1–0.2 MOD), longer and sparser on pleura and mesoscutellum (0.3–0.4 MOD). Mesoscutum with scattered, plumose, long hairs (up to 1.2 MOD), longer on pleura (1.6–2.1 MOD) and metanotum (up to 1.5 MOD). Lateral area of propodeum with plumose hairs up to 3.6 MOD. Anterior part of T1 with plumose hairs (1.2–1.5 MOD), shorter and sparser on disk. T2–T4 with simple, short (0.2 MOD) hairs, and some scattered, plumose longer hairs (0.5–1.5 MOD), directed posteriorly; apical impressed areas with simple, short hairs. Sterna glabrous basally, with long (2.7–3.5 MOD), mostly simple hairs on posterior half, those on S2– S3 with their apices bent down, posteroventrally oriented.
Sculpture. Labrum with verrucose, median, basal elevation ( Fig. 1 View FIGURE 1 F). Clypeus and supraclypeal area with punctures separated by 2.0–3.0 PD, denser on apical half of clypeus. Lower paraocular area with punctures separated by 3.0–4.0 PD; upper paraocular area with punctures separated by 0.5–1.0 PD. Punctures on disc of mesoscutum separated by 4.0–5.0 PD, denser at sides, those of the mesoscutellum separated by 1.0–3.0 PD; punctures evenly distributed. Hypoepimeral area with punctures separated by 4.0–5.0 PD, those on lateral area of propodeum separated by 5.0–7.0 PD. Metapostnotum with fine striae reaching half of length of dorsal surface in some specimens; apical margin roughly tessellate ( Fig. 1 View FIGURE 1 G). T2–T4 with punctures separated by 3.0–4.0 PD, sparser on disk of T1 (punctures separated by 5.0–6.0 PD). Surface between punctures tessellate throughout body, except terga and sterna substrigulate.
Structure. Head broader than long, 1.29–1.32:1. Ratio of lower to upper interocular distance 0.93–0.98:1. Clypeus broader than long, 1.75–1.83:1. Ratio of interantennal to antennocular distance, 0.52–0.62:1. Ratio of posterior interocellar to ocellocular distance, 0.99–1.10:1. Inner hind tibial spur with many teeth, shorter than diameter of shaft of spur ( Fig. 1 View FIGURE 1 E).
Antennal sensilla. One individual with a bare area free of sensilla on the most apical flagellomere (i.e. no sensilla occur in an oval-like area on the ventral side of F10), the other two lacking such bare area. SP intermingled with other types of sensilla all throughout all flagellomeres (i. e. no large, distinct areas of exclusive SP were observed as in the male). SP about 15–20 µm long, with longitudinal axis roughly parallel to the longitudinal axis of the antenna ( Fig. 2 View FIGURE 2 C). SP denser on the dorsal side of the antennae and on the distal flagellomeres ( Fig. 2 View FIGURE 2 D–G). SB ( Fig. 2 View FIGURE 2 H–I) with a peg 5–7 µm long, sculptured longitudinally. SB fully erect ( Fig. 2 View FIGURE 2 H) or partially curved ( Fig. 2 View FIGURE 2 I). SB rare, mostly on the ventral side of the flagellomeres ( Fig. 2 View FIGURE 2 E). SAm clearly smaller (about 0.5–1 µm) than SCo (about 1.5 µm) (peg often visible protruding partially from the pit) ( Fig. 2 View FIGURE 2 C,J–K). SAm and SCo on the ventral and sometimes the latero-ventral side of most flagellomeres, often in clusters ( Fig. 2 View FIGURE 2 C,J). ST-A: 10–20 µm long ( Fig. 2 View FIGURE 2 D), very abundant, in particular dorsally ( Fig. 2 View FIGURE 2 D). ST-B: 10–20 µm long ( Fig. 2 View FIGURE 2 E); found both ventrally and dorsally. ST-C/D: variable in size (from 15 to 40 µm long) ( Fig. 2 View FIGURE 2 L); found both ventrally and dorsally and often almost perpendicular to the antenna surface. The antennae are also covered with setae of different shapes and sizes ( Fig. 2 View FIGURE 2 F,M).
Etymology. This species is named moscosensis after Los Moscos, a lake in Nahuel Huapi National Park ( Argentina ) where the holotype was collected.
Comments. The male designated as the holotype is housed at MACN. This recently collected specimen is in good condition, and it only lacks the mid right leg from the femur, which was removed for DNA barcoding (658 bp: BOLD number HALIC159–11; GenBank accession KU983418 View Materials ). The holotype has the following labels: “ARG. Río Negro, PN Nahuel / Huapi: Lago Los Moscos / –41.346917 –71.664306 / 28–I–2011 R. González V. / Sobre Escallonia virgata ”, “MACN-En / 8207”, “Barcoding of Life / DNA voucher specimen / MACN-En 8207” and “ Corynura (Corynura) / moscosensis n. sp. / HOLOTYPE / González-Vaquero 2016” printed.
This species was mentioned as “ Corynura sp3a” in a recent molecular study (González-Vaquero et al., 2016), and it can be distinguished from 14 other Corynura species by its Barcode Index Number ( Ratnasingham & Hebert 2013): BOLD:ABW7139. Unfortunately, C. chloromelas was not included in that analysis due to lack of barcode compliant sequences of this species. Besides the holotype, three paratype females from Neuquén ( Argentina ) have been barcoded, all 658 bp sequences: MACN-En 8163 (BOLD number HALIC140–11; GenBank accession KU983415 View Materials ), MACN-En 8167 (BOLD number HALIC144–11; GenBank accession KU983416 View Materials ), and MACN-En 8168 (BOLD number HALIC145–11; GenBank accession KU983417 View Materials ). These sequences and those studied by González-Vaquero et al. (2016) are publicly available online (project CORYN, http://www.boldsystems.org).
In some male specimens, the process on the posterior margin of the S8 is difficult to observe because it is membranous and translucent. This process, which is glabrous in the holotype, may bear one or two apical setae. In the male antennae, although SB, SAm and SCo were not observed, we cannot confirm their absence in the male sex of this species due to the limited sample size (one specimen).
Distribution. In Argentina : southwestern Neuquén, western Río Negro and northwestern Chubut. In Chile: from La Araucanía to Los Lagos. This species inhabits in the Andean Nothofagus -dominated forest, in the biogeographic province of the Valdivian Forest, Subantactic sub-region ( Morrone 2015).
Examined material: Paratypes. ARGENTINA : Neuquén: 7f, Arroyo Pedregoso , 10–XII–2001, D. Medan, N.H. Montaldo, M. Devoto et al. ( FAUBA) . Lanín National Park: Boquete (Lago Lolog): 1m, 4–IV–1964, M. Gentili ( MACN) ; 1f, 4–IV–1964, M. Gentili (IADIZA); 1f, 25–IV–1964, A.J. Giai (SEMC). 1f, Lago Huechulafquen , margen N, 15–XII–2010, L. Compagnucci & R. González Vaquero (MACN-En 8163; MACN) . Lago Queñi : 1f, 16–XII–1999, N.H. Montaldo, M. Devoto & G. Gleiser ( FAUBA) ; 1f, 2–XII–1985, A. Roig Alsina (MACN); 1f, 20–I–1954, M.M. Senkute (SEMC). Nahuel Huapi National Park: Isla Victoria: 4f, Schajovskoi ( MLPA) ; 2f, X–1963 (IADIZA); 1f, X–1963, A.J. Giai (SEMC); 1f, XII–1954, R.N. & J.S. de Orfila (SEMC). 2f, Puerto Arrayán , 20–XII–2010, L. Compagnucci & R. González Vaquero (MACN-En 8167 and MACN-En 8168; MACN) ; 29f, Puerto Arrayán , 20–XII–2010, L. Compagnucci & R. González Vaquero ( MACN) . Río Negro: 1f, R.S. Castillo ( MACN) . Nahuel Huapi National Park: Puerto Blest : 2f, XII–2000, C. Morales & C. Quintero ( MACN) ; 1f, 15–IX–2001, C. Morales & C. Quintero (MACN). Chubut: INTA Trevelin : 10f, 2–XII–2005, A-I. Gravel ( MACN) ; 5f, 6–XII–2006, A-I. Gravel (MACN); 1f, 7–XI–2005, A-I. Gravel (PCYU); 1f, 20–XII–2005, A-I. Gravel (PCYU); 15f, 2–XI / 11–XII–2006, A-I. Gravel (PCYU); 12f, 8–XI / 13–XII–2006, MIK. Gravel (PCYU). Los Alerces National Park: Lago Futalaufquen : 9f, 13/ 18–XII–1997, C. & M. Vardy Malaise trap ( BMNH) ; 10f, 14–XII–1997, C. & M. Vardy (BMNH); 2f, Bahía Mansa , 17–II–2011, J.L. Farina (MMLS); 15f, 14–XII–1997, C. & M. Vardy (MACN). CHILE: Araucanía: 1f, Cabreria, Cordillera Nahuelbuta , 22–XII–1985, A. Roig Alsina ( MACN) . Los Ríos: 1f, Niebla , 14–XII–1985, A. Roig Alsina ( MACN) . Los Lagos: 2m, Guabún, W Ancud, 13–I–1981, L.E. Peña ( AMNH) . 1f 1m, El Chinque, N of Correntoso , 22–I–1980, L.E. Peña ( AMNH) . 1f, La Picada, NW Volcán Osorno, 16/ 18–I–1980, L.E. Peña (AMNH). 1f 1m, Puyehue , 10–II–1979, L.E. Peña ( AMNH) . 1m, Río Gol Gol , 8/ 13–II–1957 ( SEMC) . 2f, Termas de Puyehue , 4/ 5–XII–1985, A. Roig Alsina ( MACN) . 1f, Anticura on Río Golgo, E of Puyehue , 18/ 29–X–1985, L.E. Peña ( AMNH) . 1f 1m, Peulla , 7–II–1974, L. Ruz ( PCYU) . Hualaihué, Estación Científica Huinay , 35 m, nesting aggreg. in path, 21–X–2014, C. Polidori: 5f ( MNCN), 2f ( MACN) . 1m, Huinay , I–1981, O. Martinez ( PUCV) . 1f, Parque Nacional Puyehue, El Pionero , 16– XI–2000, L. Packer ( PCYU) .
Floral associations. The plants listed below were taken from the specimen labels, which were collected in recent field trips made by RAGV to Neuquén and Río Negro ( Argentina ): Anacardiaceae : Schinus patagonicus (Phil.) I.M.Johnst. ; Asteraceae : Baccharis obovata Hook. et Arn. ; Boraginaceae : Phacelia secunda J.F. Gmel. ; Elaeocarpaceae : Aristotelia chilensis (Molina) Stuntz ; Escalloniaceae : Escallonia virgata (Ruiz & Parv.) Pers. ; Fabaceae : Cytisus scoparius (L.) Link, Trifolium repens L., Vicia nigricans Hook. & Arn. ; Grossulariaceae : Ribes magellanicum Poir ; Malvaceae : Tilia moltkei L.; Proteaceae : Embothrium coccineum J.R.Forst. & G.Forst , Lomatia hirsuta Diels ex J.F.Macbr. ; Rhamnaceae : Discaria articulate (Phil.) Miers , D. chacaye (G. Don) Tortosa ; Rosaceae : Rosa rubiginosa L..
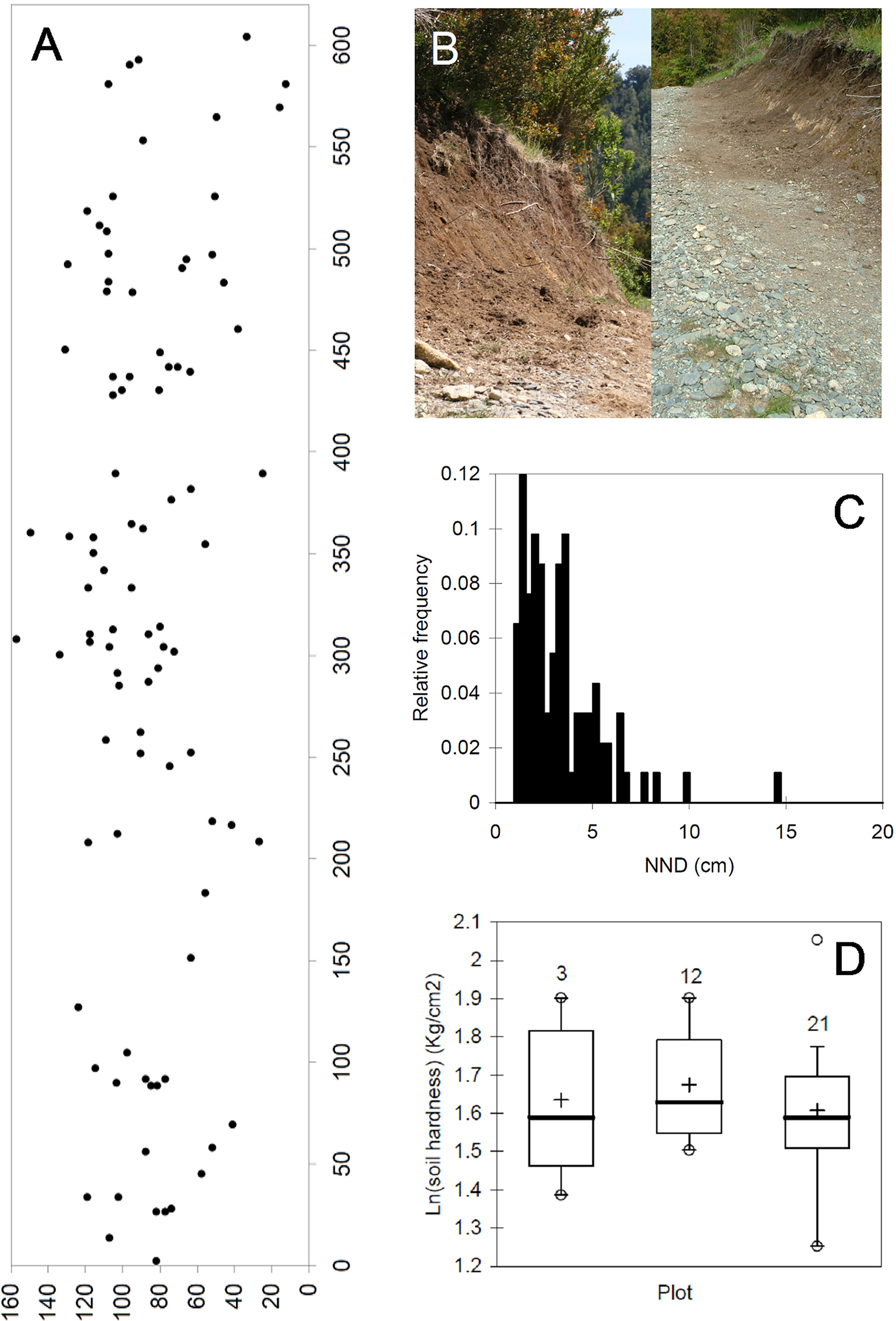
Nesting biology. The nesting site of C. moscosensis n. sp. consisted of NW-facing earth bank 15.0 m long and 1.6 m tall at the border of a ground path ( Fig. 3 View FIGURE 3 B). The bank was bordered by flowering fields with many shrubs mainly belonging to Myrtaceae , and some Nothofagus spp. and Eucryphia cordifolia Cav. trees. The bee nests (92 in total) were found in a smaller area of 6.0 m × 1.6 m ( Fig. 3 View FIGURE 3 A) rather than throughout the bank. Nest entrances lacked soil turrets.
Nest density in the nesting area was 9.6/m2. Mean distance between pairs of nests was 167.8 ± 141.2 cm, but nests’ nearest neighbours (NND) were only 3.3± 2.1 cm (range: 1.2–14.7 cm), with most NND falling between 1.0 and 3.5 cm ( Fig. 3 View FIGURE 3 C). The Clark and Evans’ test (1954) revealed a significantly clumped distribution pattern of nests (R = 17.07, n = 92; C = –13.65, P <0.001). Soil hardness overall ranged between 2.5 and 6.8 kg /cm2, and did not differ between areas with low, moderate or high nest density (ANOVA, F = 0.31, df = 2,29, P = 0.73) ( Fig. 3 View FIGURE 3 D).
Foraging activity in 2013 was low during the study, most probably because many days were cloudy, with temperature often below 20 °C, and with some rain. About 5–15 bees were observed returning to their nests with pollen on their scopae in a 4-hour period in sunny or partially cloudy days. The color of pollen brought to the nests ranged from yellow through various tones of orange to fully red. Some bees were observed in the morning or in general under cloudy and cold weather with the head visible at the entrance, possibly waiting for adequate conditions to fly or in a nest-guarding behaviour ( Fig. 4A View FIGURE 4 ). When exiting from the nests, females sometimes spent a few minutes walking on the ground, presumably to thermoregulate before flying ( Fig. 4 View FIGURE 4 B). In 2014 no bees were observed out of the nests due to the frequent and strong rainfalls which occurred on almost every day of the study.
In 2013, during nest excavations of an area including 11 nest entrances, a total of 37 brood cells were discovered. These cells were ovoid in shape and were located at 20–40 cm in depth, and at about 5–10 cm from a central tunnel that descended obliquely from the surface. We were unable to find clear clusters of cells, though sometimes cells were quite close to one another. Although we could not get information for individual nests, given the number of entrances in the excavated area we can infer that the minimum number of cells per nest may be around three at this early stage of nest development. Two of the brood cells found contained pollen/nectar stores (yellowish in color). An additional 15 cells were empty but apparently lined ( Fig. 4 View FIGURE 4 C), without any faeces and thus possibly new cells to be provisioned, while four cells contained an adult alive bee ( Fig. 4 View FIGURE 4 D), with unworn or weakly worn wings ( Fig. 4 View FIGURE 4 D). Two other cells included puparia of Diptera , most probably Leucophora spp. ( Diptera : Anthomyiidae ). Many Leucophora flies were observed performing their typical “satellite flights” (i.e. they reach the bee nests by following the bees returning to the nest in flight) and were determined as L. andicola (Bigot) and L. peullae (Malloch)) ( Polidori et al. 2015) ( Fig. 4 View FIGURE 4 E). The remaining 10 cells appeared to be old, with remains of faeces and partially filled with soil. In that year, 10 females of C. moscosensis n. sp. were collected from the nest excavations and four females were collected while leaving their nests. In 2014, nest excavations of an area that included 22 nest entrances provided 18 additional females. In that year, the soil was wet and muddy, making the recognition of tunnels and brood cells difficult.
The ratio between the number of bees found in the tunnels and the number of nest entrances in the excavated area was 0.9 in the middle of the spring and 0.8 in late spring. In 2013, three females were found together in two tunnels and two females in another one, while the other bees were individually found within indistinguishable tunnels or in brood cells.
While we did not found immatures in the bee nests, a further natural enemy of this bee at this locality may be the velvet ant Dimorphomutilla suavissima (Gerstaecker) ( Hymenoptera : Mutillidae ) ( Fig. 4 View FIGURE 4 F), because females of this species were found walking on the nesting area and inspecting nest tunnels, particularly in the morning.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |