
Xetadrilus jejuensis, Dózsa-Farkas & Felföldi & Nagy & Hong, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4496.1.27 |
|
publication LSID |
lsid:zoobank.org:pub:7C536E1E-5D5A-4E2D-9E4F-28F3CEA9664C |
|
DOI |
https://doi.org/10.5281/zenodo.5950213 |
|
persistent identifier |
https://treatment.plazi.org/id/03D3D43A-E45E-FFB5-2580-F9DAFDCDFD2F |
|
treatment provided by |
Plazi |
|
scientific name |
Xetadrilus jejuensis |
| status |
sp. nov. |
Xetadrilus jejuensis View in CoL sp. n.
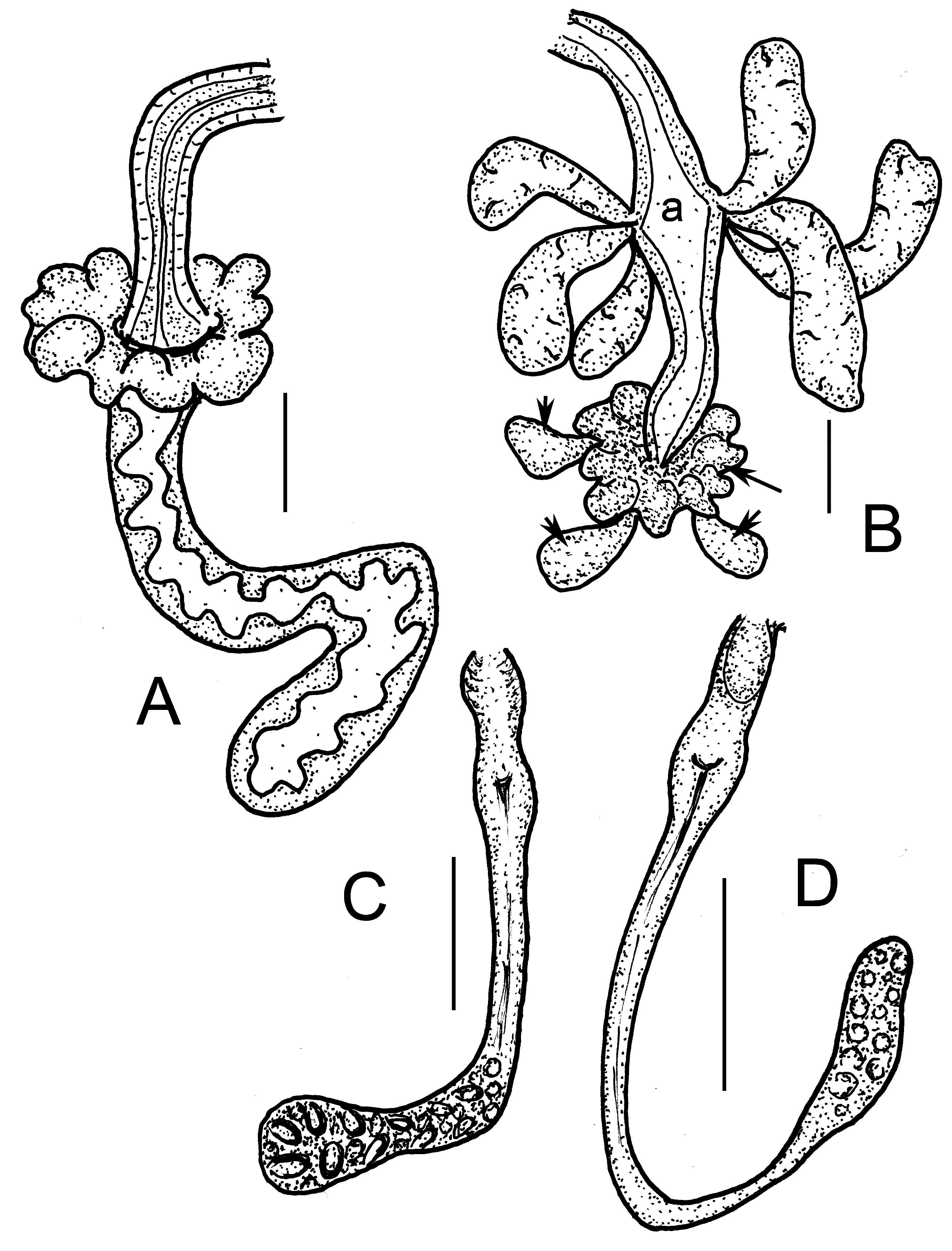
( Figures 12D View FIGURE 12 , 15–16 View FIGURE 15 View FIGURE 16 )
Type material. Holotype: NIBRIV0000810596, slide No. 2206, adult, stained whole mounted specimen. Type locality site 7, Mt. Hallasan in Gwaneumsa trail, Jeju Island, Korea, soil and litter layers of Styrax japonica tree, N 33˚23'15.9", E 126˚32'03.3", 1306 m asl, 0 9.06.2016, leg. Y. Hong. Paratypes (in total 13 stained, adult specimens on slides and 34 specimens in 70% ethanol): NIBRIV0000810597, slide No. 2176, site 8, NIBRIV0000811386, slite No. 2396, site 8, P.119.1.1–119.1.8, slide No. 2203–2205, 2207, 2399–2402, from type locality, P.119.2.1– 119.2.2, slide No. 2397–2398, site 8, P.119.3, slide No. 2300 (last 6 segments, 0.8 mm for DNA analysis No. 1129). In 70% ethanol: P.119.4, twenty-three specimens, type locality; P.119.5 site 8 one specimen; P.119.6, site 6 one specimen; P.119.7, site 11 five specimens; P.119.8 site 12 four specimens.
Further material examined: 10 specimens, only in vivo.
Etymology. Named after Jeju Island.
Diagnosis. The new species can be recognized by the following combination of characters: (1) small worms, 2.5–4.5 mm long, width about 130–200 µm in vivo, segments 23–31; (2) lateral chaetae present in II–VII, absent from VIII on, formula: 2,0–0: 2–2; (3) clitellum saddle-shaped: dorsally granulocytes and hyalocytes, laterally only granulocytes; (4) in prostomium at the frontal tip present a vesicle-like recess, 6–8 inner papillae and a pair of prostomial ganglia; (5) brain incised posteriorly; (6) dorsal blood vessel from XII–XIII, blood colourless; (7) pharyngeal gland with unpaired dorsal lobes in IV and V, secondary ventral lobes in V and VI spherical, in VI–VII a pair of separate elongate lobes with a “Z-like fashion”; (8) two pairs of preclitellar nephridia in 6/7– 7/8; (9) sperm funnels 60–90 µm long in vivo, 2–3 times longer than wide, collar distinct; (10) male copulatory organs small, spherical glandular bulbs diameter 35–40 µm in vivo, extra bulbs absent; (11) spermathecae free, at the orifice of ectal ducts with a sessile gland, distally widening into a narrow ampulla, the following tubes widening into thin-walled ental reservoirs in VII or VIII.
Description. Small, slender worms ( Fig. 15A View FIGURE 15 ). Holotype 2.5 mm long, 100 µm wide at VIII and 150 µm at clitellum (fixed), 23 segments. Length of paratypes 2.5–4.5 mm, width 130–180 µm at VIII and 170–200 µm at clitellum in vivo, length of fixed specimens 2.5–4 mm, width 100–150 µm at VIII and 150–190 µm at clitellum, segments 23–31. Chaetae two per bundle ( Fig. 15D View FIGURE 15 ), straight, slightly bent proximally 17–23 µm long and 2 µm thick preclitellarly, at the hindmost segments about 33–40 µm long and 2 µm thick. Lateral chaetae present in II– VII, absent from VIII on. Ventral chaetae from II on, absent in XII, formula 2,0–0: 2–2. On the body surface pale elongate glandular cells in 7–8 transverse rows and some roundish gland cells. Clitellum XII–1 /2XIII, saddleshaped, not developed ventrally, the gland cells in about 18 transverse rows, dorsally granulocytes and hyalocytes but laterally and downwards towards the male copulatory organs only granulocytes ( Fig. 15E View FIGURE 15 ). Prostomium similar to other Xetadrilus species, the head pore in mid-dorsal position on the prostomium ( Fig. 15B View FIGURE 15 ). Frontal prostomial epithelium thickened, with a vesicle-like recess at the frontal tip. About 6–8 inner papillae and a pair of prostomial ganglions (15–22 µm long, fixed) present in prostomium ( Fig. 15B View FIGURE 15 ). Prostomial musculature well developed. Body wall about 12–15 µm and cuticle 1–1.5 µm thick in vivo, septa 4/5– 9/10 thickened.
* according to Schmelz et al. 2011
Brain about 90–100 µm long, 2 times longer than wide (fixed), incised posteriorly ( Fig. 15C View FIGURE 15 ). Post-pharyngeal bulbs well developed. Perikarya of II–IV fused into suboesophageal ganglion, segmental ganglia present from V on ( Fig. 15B View FIGURE 15 ). Pharyngeal glands ( Figs. 15F View FIGURE 15 , 16G–H View FIGURE 16 ) with unpaired dorsal lobes in IV and V, primary ventral lobes in V elongate; secondary ventral lobes in V and VI spherical, smaller than primary ventral lobes. In VI–VII a pair of separate elongate lobes, consisting of an anterior dorsal part in VI and a posterior ventral part in VI–VII; both parts broadly connected in Z-like fashion ( Fig. 16G–H View FIGURE 16 ). Oesophageal appendages and intestinal diverticula absent, intestine widening abruptly in VII. Chloragocytes small, light brown, about 10–13 µm long in vivo. Dorsal vessel from XII–XIII, blood colourless. Midgut pars tumida from XVIII–XXI, extending over 3 segment lengths ( Fig. 16C View FIGURE 16 ). Two pairs of preclitellar nephridia from 7/8 to 8/9, not constricted at septums; anteseptale as long as postseptale or slightly shorter, terminal origin of efferent ducts ( Fig. 15G View FIGURE 15 ), dorsal vesicle in the postseptale absent. First postclitellar nephridia at 13/14. Coelomocytes broadly oval, ca. 16–23 µm long, distinctly dark brown in vivo, filled with small refractile vesicle ( Fig. 16D,E View FIGURE 16 ), in fixed specimens the cytoplasma pale with well visible nucleus, 12–19 µm long ( Fig. 16A View FIGURE 16 ). Seminal vesicle absent. Sperm funnels 60–90 µm long in vivo 2–3 times longer than wide (50–80 µm long and 2 times longer than wide, fixed), tapering distad, collar distinct, about as wide as funnel body or slightly narrower ( Figs. 15E View FIGURE 15 , 16D–F View FIGURE 16 ). Sperm ducts 6 µm wide. Length of spermatozoa 30–50 µm, heads 12–15 µm in vivo (30–40 and 7–10 µm, fixed). Male copulatory organs ( Fig. 16B View FIGURE 16 ) small, spherical glandular bulbs diameter 35–40 µm in vivo (25–30 µm, fixed) extra bulbs absent. Spermathecae ( Figs. 12D View FIGURE 12 , 16G–H View FIGURE 16 ) not attached to oesophagus. Ectal ducts short (length 20–30 µm, diameter 12–15 µm in vivo) at the orifice with a sessile gland ( Fig. 16I View FIGURE 16 ); distally widening into ampullae (diameter 16–17 µm), from here sperm present, which is arranged in parallel in longitudinal axis of spermatheca. The following tube narrower (diameter ca. 9 µm, 150–200 µm long in vivo), widening into thin-walled ental reservoir in VII or VIII (sometimes bent backwards into VI). The reservoir-sacks about 40–100 µm long and 10–20 µm wide in vivo and mostly filled with spheroid bodies ( Fig. 16G View FIGURE 16 ). Mostly one large mature egg at a time.
Distribution and habitat. Korea, Mt. Hallasan, Jeju Island, site 6–8, 10–12: soil and litter layers under Quercus mongolica , Styrax japonica , Quercus serrata , Sorbus alnifolia and mixed forest.
Differential diagnosis. Up to now, three Xetadrilus species have been reported with dark coelomocytes: X. pitucus ( Righi, 1974) , X. righianus ( Xie & Rota, 2001) and X. maacki Schmelz, Collado & Römbke, 2011 . But in X. pitucus , all three pharyngeal glands are united dorsally, the secondary lobes are absent and the lateral chaetae are absent already from VII. In X. righianus , the secondary lobes of pharyngeal glands also absent, the clitellar glands occur also ventrally but are weakly developed and the spermathecal reservoir is globular (diameter 37–40 µm). In X. maacki , all pharyngeal glands are united dorsally, three ventral pairs of secondary lobes are present and the spermathecae are confined to V. Similar and distinguishing traits of Xetadrilus jejuensis sp.n., X. aphanoides sp. n. and X. aphanus are listed in Table 2.


| DNA |
Department of Natural Resources, Environment, The Arts and Sport |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Oligochaeta |
|
Order |
|
|
Family |
|
|
Genus |