
Williamsium parviflorum ( Thomson, 1916 ), 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4304.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3D557C94-0783-4C39-80C3-9C321DA94800 |
|
DOI |
https://doi.org/10.5281/zenodo.6015379 |
|
persistent identifier |
https://treatment.plazi.org/id/039B87ED-3E36-FF30-FF4B-E37E783EDB32 |
|
treatment provided by |
Plazi |
|
scientific name |
Williamsium parviflorum ( Thomson, 1916 ) |
| status |
comb. nov. |
Williamsium parviflorum ( Thomson, 1916) View in CoL new combination
( Figs. 90–97 View FIGURE 90 View FIGURE 91 View FIGURE 92 View FIGURE 93 View FIGURE 94 View FIGURE 95 View FIGURE 96 View FIGURE 97 )
Anthothela parviflora Thomson, 1916: 3 View in CoL –6, Pl. II Fig. 5 View FIGURE 5 , Pl. V Fig. 4 View FIGURE 4 ; Williams 1992a, 185–186, Figs. 2–3 View FIGURE 2 View FIGURE 3 , 4 View FIGURE 4 A–D.
Material examined. Paralectotype: NHMUK 1962.7 About NHMUK .20.40, off Algoa Bay , South Africa, ‘P.F. 524’, depth 183 m, 1st November 1898, S.J. Hickson collection.
Description:
Colony form: The fragments examined here are from one of three syntypes, which was only briefly mentioned in Thomson’s (1916) original description. His description was based on the “most complete example” which was probably the specimen designated as the lectotype by Williams (1992a). Unfortunately it was not possible to view the lectotype which is stored at the South African Museum. The material examined here is from a different location to the lectotype but corresponds with Thomson’s extensive description and strongly resembles the figures in Williams (1992a).
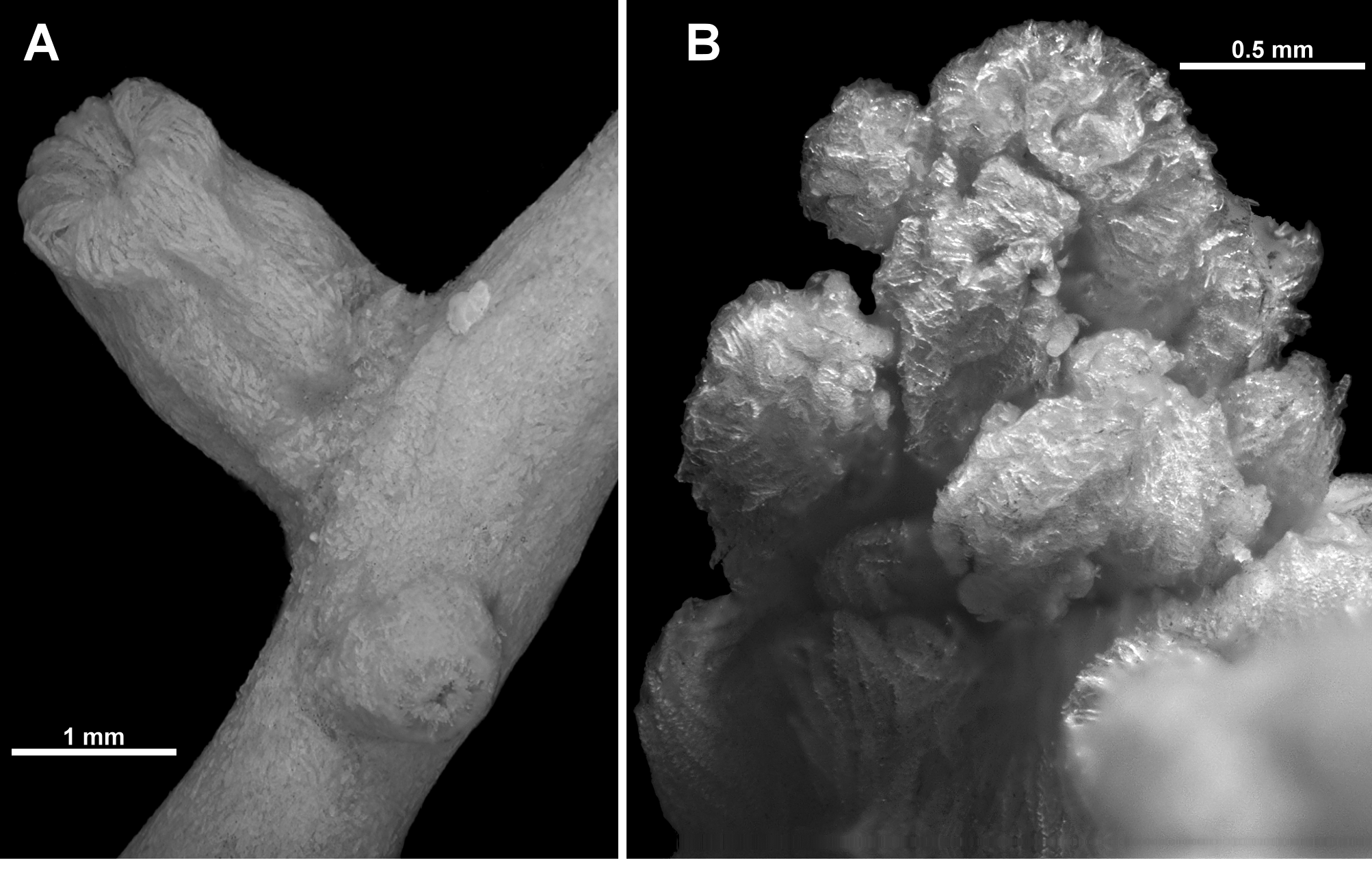
The complete lot consists of a small, incomplete colony plus the three fragments examined here ( Fig. 90 View FIGURE 90 A). They are all pieces of branches with tall calyces emanating at right angles or obliquely from the branch ( Fig. 90 View FIGURE 90 B). The largest fragment is 51.5 mm long with eleven polyps spread evenly along it; another fragment is 28 mm long with seven visible polyps (approximately half of the branch is surrounded by an encrusting sponge); and the smallest fragment is 14 mm long with four polyps. The two smallest pieces have intact branch tips which have small clumps of adult polyps (one has two juvenile polyps on the very apex of the branch). None of the fragments examined have any evidence of branching, however the largest portion of the colony pictured in Fig. 90 View FIGURE 90 A is consistent with the species description by Thomson; that is, colonies with a spreading base, sparse, irregular branching, basically in one plane, bifurcation at approximately 45 degrees and branches which are twisted or curved. There are no anastomoses noted for the three specimens described by Thomson (1916) or the four colonies examined by Williams (1992a), although Thomson does mention that colonies can be “creeping”, perhaps indicating this species can grow both membranous and branching forms. Williams, however, does not mention any such growth form from the four “whole” colonies included in his description.
The three branch fragments are circular in cross-section and diameter varies little (from 1–2 mm). In the larger colony portion pictured, the major branches are up to 3 mm wide while the bulky basal stem is approximately 5 mm wide.
All fragments are in good condition with mostly intact polyps and an undamaged colony surface.
Colour: No mention is made of live colour by either Thomson or Williams, although Thomson mentions that the (presumably preserved) colony has a “slightly silvery appearance”. This is more likely due to the dense layer of sclerites than colony colour. The fragments are now cream in alcohol.
Polyps and calyces: Calyces are sparsely arranged on all sides of the branches, approximately 3.5–5 mm apart. Terminal clumps of calyces are not large or overly crowded ( Fig. 90 View FIGURE 90 C). The calyces are tall and cylindrical, with eight distinct longitudinal furrows. Most calyces are approximately 3–4.5 mm tall and 1.5–2.0 mm wide although there are rare calyces only 2–2.5 mm tall. The exsert part of the polyps are approximately 1–2.3 mm tall but none are fully extended—most have the polyp head resting on the lip of the calyx ( Fig. 90 View FIGURE 90 D). Polyp heads are approximately 1.3–1.6 mm in diameter and some are fully retracted into the calyx ( Fig. 91 View FIGURE 91 A). The tentacles are often crumpled in a haphazard way over the polyp mouth ( Fig. 91 View FIGURE 91 B), although there are occasionally polyps with the tentacles folded in, across the polyp mouth, giving those polyps a mounded, starred apex to the polyp head. There is a single row of 10 very long and narrow pinnules along each side of the tentacles. The pinnules taper to a sharp tip and often twist and curl.
Medulla and Cortex: The medulla is constructed of tightly packed sclerites, arranged longitudinally in general, and is surrounded by a cortex, similarly consisting of crowded sclerites. The cortex is distinctly separated from the medulla by a ring of relatively large and well-defined boundary canals ( Fig. 92 View FIGURE 92 A). These canals do not obviously anastomose to form a boundary space but run longitudinally, adjacent to each other, the length of the fragments. This is similar to that seen in specimens of Anthothela although here the canals are much larger relative to the diameter of the branches; within the 1–2 mm branch diameter the boundary canals have approximate diameters of 0.1–0.15 mm. The cortex is approximately 0.1–0.2 mm thick while the medulla is approximately 1-1.5 mm in diameter. There are no coelenteric canals within the medulla. Thomson (1916) mentions “a few small canals” in the medulla but there was no indication of such found here.
Due to the scarcity of material, no investigation was conducted on the canal arrangement at the branch tips.
Sclerites: Sclerites cover the calyces and polyps. On the polyp head, straight or very slightly curved, tuberculate sticks and spindles are arranged in eight points ( Fig. 92 View FIGURE 92 B, C). These sclerites are 0.18–0.38 mm long and are arranged longitudinally on the central ridge of the points, and en chevron to obliquely on the sides of the points. A collaret is not present. There are intermediate sclerites arranged longitudinally between the points, with clumps of up to five proximally, reducing to only one or two distally ( Fig. 92 View FIGURE 92 Ba). They are similar in form to the sclerites of the points. In the distal part of the points, the longitudinally arranged sticks and spindles cease and are replaced by transversely arranged, small, lightly tuberculate, mostly flattened rods ( Fig. 93 View FIGURE 93 A, B) along the back and side of the rachis. They continue to the tip of the tentacle, decreasing in size distad, and range from 0.11–0.21 mm in length. The pinnules have numerous small, very lightly tuberculated scales ( Fig. 93 View FIGURE 93 C). Some are butterflyshaped or have a waist but most or straight with slightly crenulated edges. They are arranged transversely in the pinnules ( Fig. 93 View FIGURE 93 A), and although numerous, are not overly crowded. Their length ranges from 0.05–0.12 mm.
The polyps have an irregular arrangement of sclerites spread over the neck region, mostly short rods with simple tubercles ( Fig. 94 View FIGURE 94 A). Sizes range from 0.07–0.19 mm long.
The pharynx is fleshy and thick, and when contracted shows rounded, transverse ridges which are possible muscle bands ( Fig. 94 View FIGURE 94 B). Sclerites are rare or absent proximally but distally are arranged in indistinct longitudinal groups. They are short rods with few tubercles ( Fig. 94 View FIGURE 94 C), similar to those in the neck, and differ slightly from those pharynx sclerites found in the other genera included herein. They are approximately 0.08–0.13 mm long.
On the calyces, the sclerites are arranged mostly longitudinally or slightly obliquely and are quite crowded together. They are visible as a silvery layer with individual, large sclerites discernible for most of the calyx, but at the lip they overlap so only the sclerite tips are visible. Almost all are narrow to stout spindles with simple to relatively crowded tubercles ( Fig. 95 View FIGURE 95 ) and they range from 0.06–0.32 mm long.
In the cortex, the sclerites are similar to those from the calyx, that is, sticks and spindles with simple tubercles through to complex warts ( Fig. 96 View FIGURE 96 ) and they are 0.06–0.34 mm long. There tends to be a higher percentage of warty spindles, particularly short, stout ones ( Fig. 96 View FIGURE 96 a), in the surface than in the calyx but this may be an artefact of sampling or patchiness.
The medulla contains similar warty spindles as found in the cortex, as well as simple, lightly tuberculate sticks and spindles, short, mostly smooth spindles and rare warty crosses ( Fig. 97 View FIGURE 97 ). The tuberculate sticks and spindles are easily damaged or broken during the sampling process so maximum length is an underestimate, but they appear to be 0.11–0.4 mm long. The warty spindles are relatively consistent in length ( 0.18–0.25 mm) while the mostly smooth spindles are only 0.06–0.1 mm long.
All sclerites are universally transparent and colourless under transmitted light.
Distribution: This species has rarely been reported and it is assumed it is restricted to the waters around South Africa ( Williams 1992b). Considering the report by Williams (1992a) on four full colonies in one trawl in an area outside the type locality, the species may be locally common.
Depth: 180–500 metres.
Remarks: This species was originally placed in the genus Anthothela by Thomson on the then justifiable grounds of a similar scleraxonian medulla and mostly comparable polyp and sclerite form. Based on the redefinition of Anthothela herein, this species can no longer be considered an example of the genus due to the distinct differences in the sclerites of the tentacles and pinnules and in the colony form. Anthothela species have tuberculate sticks and spindles arranged longitudinally along the rachis of the tentacle, while W. parviflorum n. comb. has short, flat rods arranged transversely along the tentacle. Similarly, Anthothela species have long, narrow-handled spatulate clubs crowded longitudinally in the pinnules where W. parviflorum n. comb. has short scales arranged transversely. Differences in colony form are also noteworthy—specimens of Anthothela have no single trunk or main stem, having instead a tangled, anastomosing colony form with little consistent structure and crowded polyps, while specimens of W. parviflorum have single trunks with no noted examples of anastomoses and only sparse branching and relatively isolated polyps.
In his discussion comparing his new species with Anthothela grandiflora, Thomson (1916) mentioned that A. parviflora has “long, thin spindles or rods with few processes” in the tentacles and that these are similar to those found in A. grandiflora “but apparently in some cases at least are much longer”. This description does not correspond to the short, flat transverse rods found in the tentacles in this study. In his re-description of the species where the lectotype was designated, Williams (1992a) does not document the placement of sclerites on the polyp, mentioning only that there are “numerous needle-like spindles or a few stout rods”. Just before publication there was an opportunity to examine the lectotype (South African Museum) and there was good congruence between the fragment examined and the paralectotype described and figured here. Thus it appears likely that Thomson was erroneously referring to the sclerites from the points.
The combination of a sparsely branching, arborescent colony with no coelenteric canals in the central medulla, widely dispersed, tall calyces, short, flat, transverse rods and scales in the tentacles and a predominance of broad, warty spindles in the calyx distinguishes this colony from all other genera in the family Anthothelidae .











| NHMUK |
Natural History Museum, London |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Octocorallia |
|
Order |
|
|
SubOrder |
Scleraxonia |
|
Family |
|
|
Genus |
Williamsium parviflorum ( Thomson, 1916 )
| Moore, Kirrily M., Alderslade, Philip & Miller, Karen J. 2017 |
Anthothela parviflora
| Thomson 1916: 3 |