
Victorgorgia argentea ( Studer, 1894 ) Moore & Alderslade & Miller, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4304.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3D557C94-0783-4C39-80C3-9C321DA94800 |
|
DOI |
https://doi.org/10.5281/zenodo.6015387 |
|
persistent identifier |
https://treatment.plazi.org/id/41B6C1CD-DF5B-4423-B857-15ECB327777C |
|
taxon LSID |
lsid:zoobank.org:act:41B6C1CD-DF5B-4423-B857-15ECB327777C |
|
treatment provided by |
Plazi |
|
scientific name |
Victorgorgia argentea ( Studer, 1894 ) |
| status |
comb. nov. |
Victorgorgia argentea ( Studer, 1894) new combination
( Figs. 110–119 View FIGURE 110 View FIGURE 111 View FIGURE 112 View FIGURE 113 View FIGURE 114 View FIGURE 115 View FIGURE 116 View FIGURE 117 View FIGURE 118 View FIGURE 119 )
Anthothela argentea Studer, 1894: 60 .
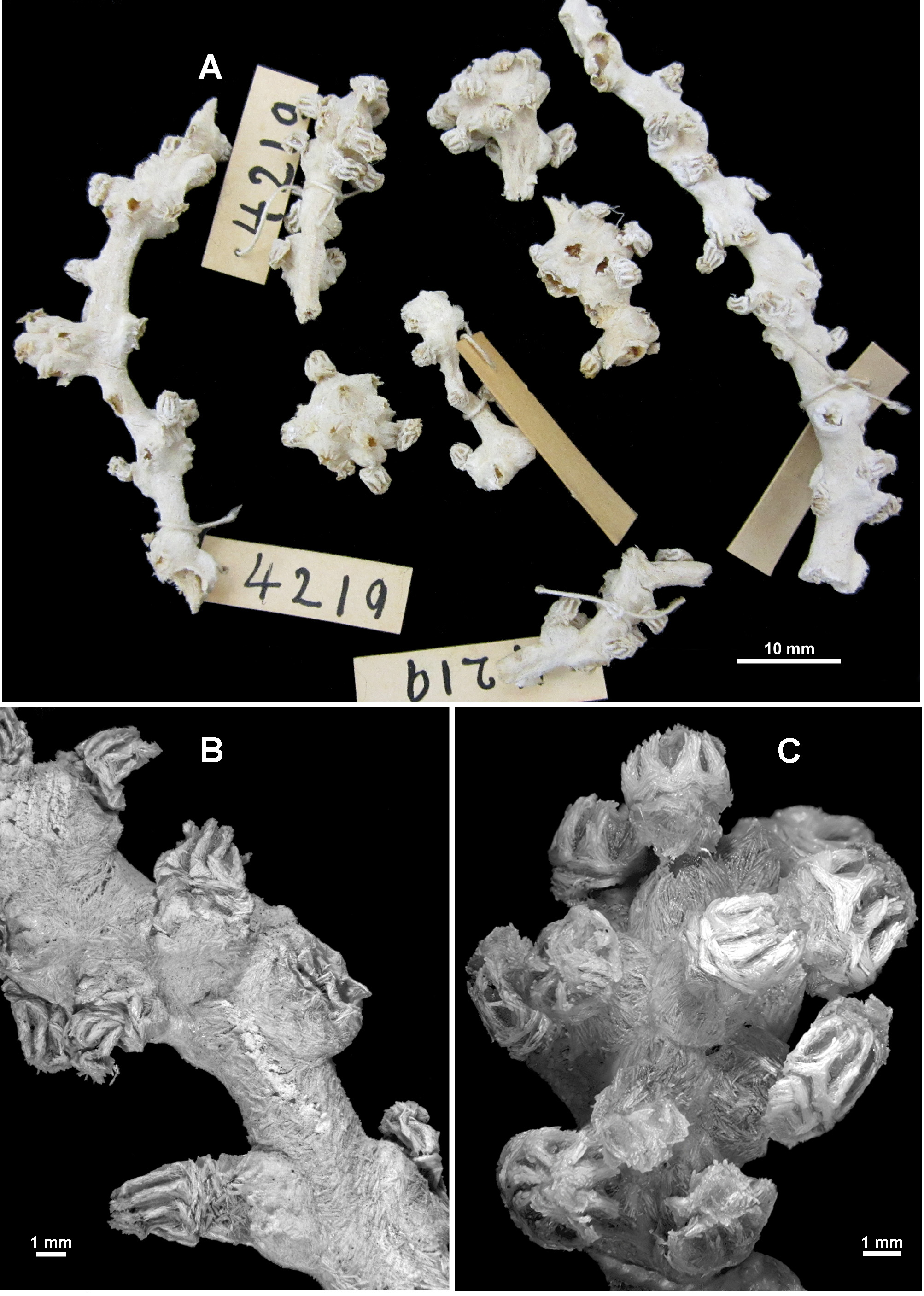
Material examined. Holotype: MCZ 4219 About MCZ , off the west coast of Mexico, U.S. Fish Commission Steamer ‘ Albatross’, stn. 3430, 23.267°N, 107.517°W, depth 1559 m, 19th April 1891. GoogleMaps
Other material: MCZ 51046, Off Oahu, 20km W of Makaha , Hawaii, T.A. Clarke collection, depth 1200 m, 5th Sept 1977.
Description:
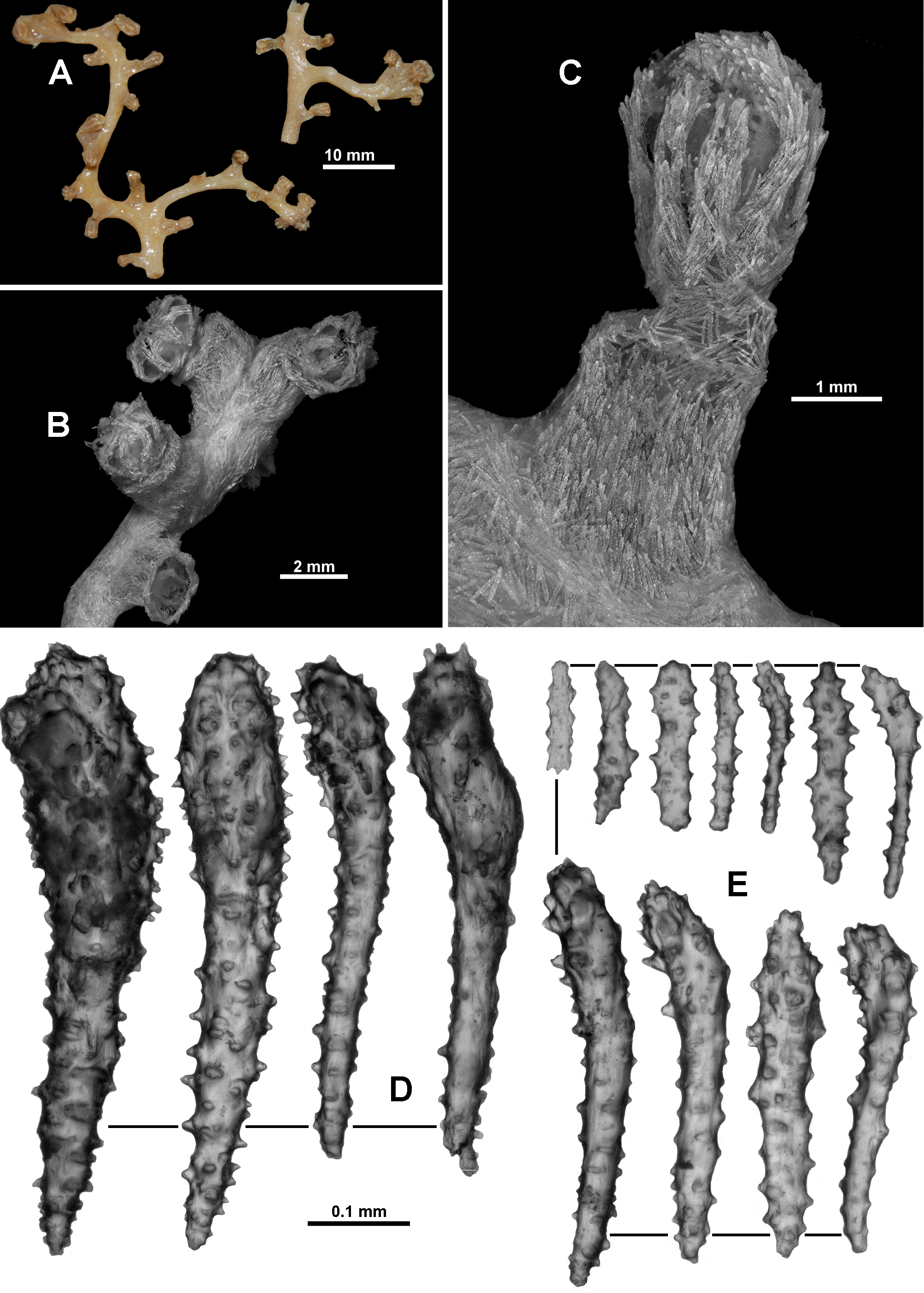
Colony form: The colony was originally described by Studer (1894) as having a “forme arborescente, les branches naissent d’un tronc principal sous des angles presque droits”, translated as having the form of a tree, with branches arising from one main trunk at almost right angles. However, the dried holotype is now in 8 recognisable pieces ( Fig. 110 View FIGURE 110 A) and numerous small fragments. Two of the largest pieces are straight portions of branch with polyps scattered along their length. One of these is 66 mm long with a diameter of 3.4 mm at the widest point where it is slightly flattened ( Fig. 110 View FIGURE 110 B). For most of the length, the branch is basically circular in cross section (approximately 2.6 mm diameter) although polyp bunches tend to cause some distortion. There is no evidence of side branches. The second fragment of branch is 62.7 mm long with a diameter of 4.2 mm at the widest point and 3.1 mm at the narrowest. There is evidence that there were two side branches on this piece, both of which have broken off at their base. Both ends of these two fragments are broken and they cannot conclusively be placed to form a larger branch. There is no evidence of anastomoses. Most of the other fragments of the holotype are terminal bunches of crowded polyps. The colony fragments and the many attached polyps are in reasonable condition although extremely brittle. There are also a number of loose polyps. A small piece of the holotype was re-hydrated for this project.
Calyces with exsert polyps are tightly crowded in bunches at the branch tip ( Fig. 110 View FIGURE 110 C) as well as being distributed irregularly along and all the way around, and generally at right angles to the branches. The largest distance between calyces is approximately 7 mm but more commonly they are closer together, in some sections touching. Occasionally, there are isolated calyces with polyps.
Colour: In the original description, Studer noted the colour of the colony to be white in alcohol, as it is now for the dried fragments. There is no record of the live colour of the colony, however Studer indicated that the large sclerites on the tentacle rachis were glassy and silvery in colour against the background brown hue of the tentacles and the re-hydrated fragment fits this description. Some of the sclerites are brown and fibrous when magnified under transmitted light, which combined with the silvery appearance, indicated the original preservation was in an acidic media, probably formalin.
Polyps and calyces: Calyces are flat-topped, conical-shaped and range in height from 1.5–2.4 mm. The polyps are relatively large, extending up to 2.5 mm from the calyx with a head diameter of between 2.2–3.4 mm ( Fig. 111 View FIGURE 111 A). Sclerites on the calyx are arranged en chevron at the base becoming longitudinal towards the lip. Most of the polyps are exsert, although often contorted or bent, likely attributable to jar storage. No polyps are fully retracted into the calyces, although some are retracted to such an extent that the polyp head sits on the top lip of the calyx and the polyp neck is hidden. The tentacles in some polyps are extended but bunched ( Fig. 111 View FIGURE 111 B) while others have the tentacles folded over the mouth such that the top of the polyps is an eight–lobed mound ( Fig. 111 View FIGURE 111 C). There are approximately 8–10 pinnules arranged in a single row along each side of the tentacles.
Medulla and Cortex: The fragile, brittle nature of the dry holotype prohibited the dissection of the branches to fully investigate the arrangement of the internal coelenteric canals. However, the broken ends of the colony pieces allow confirmation of the presence of a central medulla, consisting of tightly packed, mainly longitudinally arranged sclerites, surrounded by a narrow cortex, similarly consisting of crowded sclerites ( Fig. 111 View FIGURE 111 D, E). A boundary space appears to separate the thin, loosely attached cortex from the medulla although the fragile nature of the dried and rehydrated fragments makes this difficult to confirm. The medulla has two or three large and obvious canals (0.3–0.5 mm diameter) positioned approximately in the centre ( Fig. 111 View FIGURE 111 D, E). These significant canals are obvious at the proximal end of the colony fragments as well as just below the polyp bunches on the ends of the branches, suggesting they extend throughout the colony.
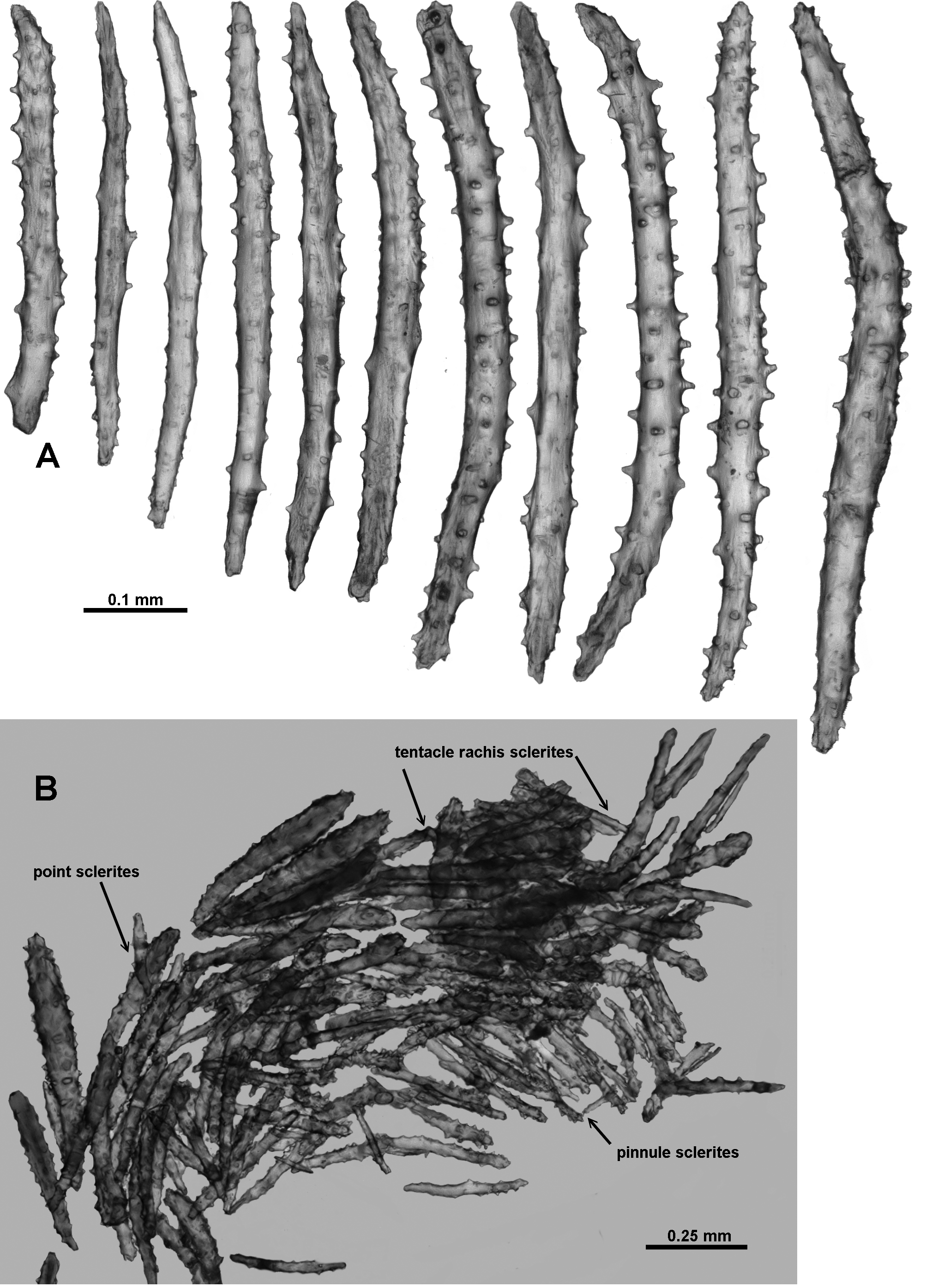
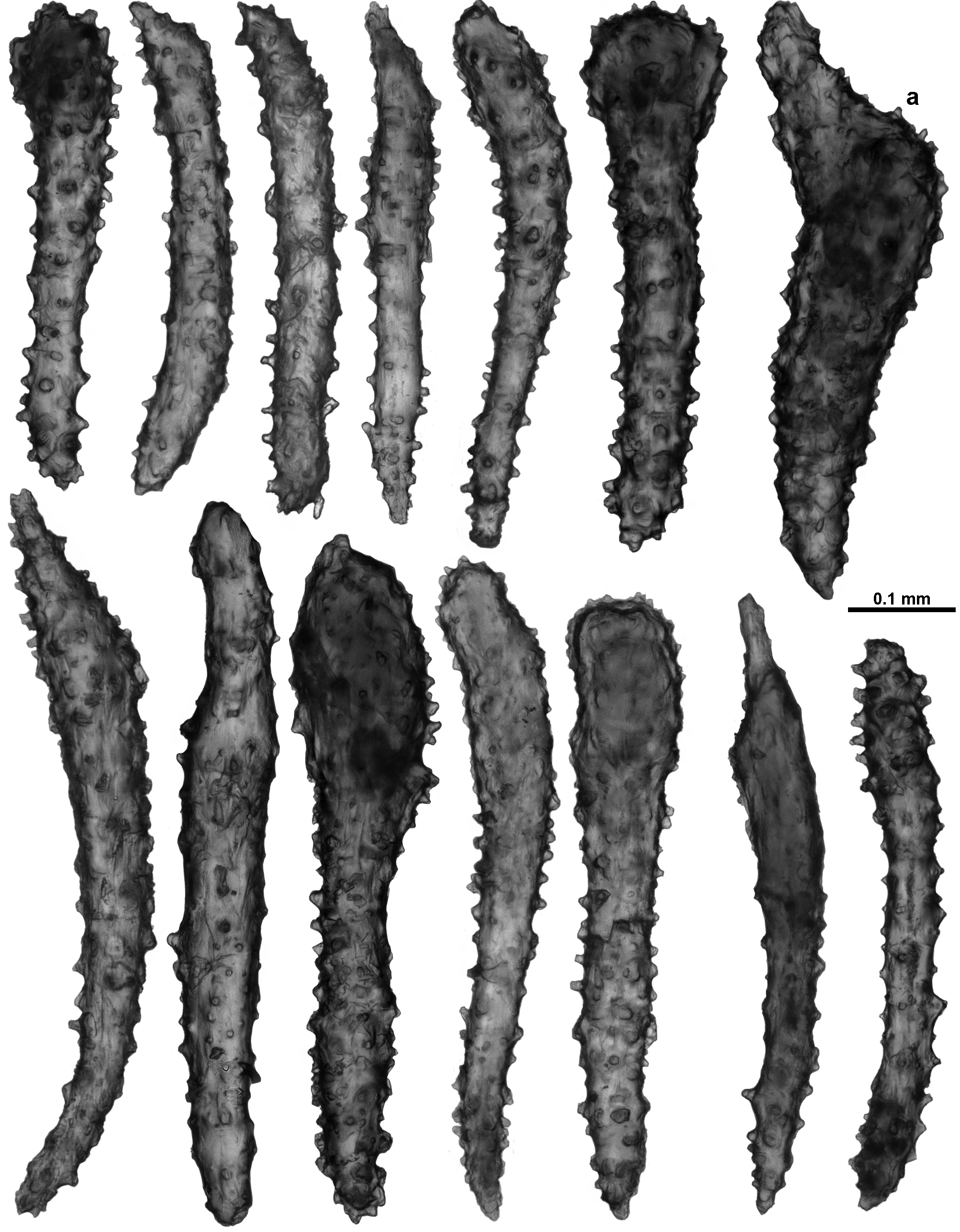
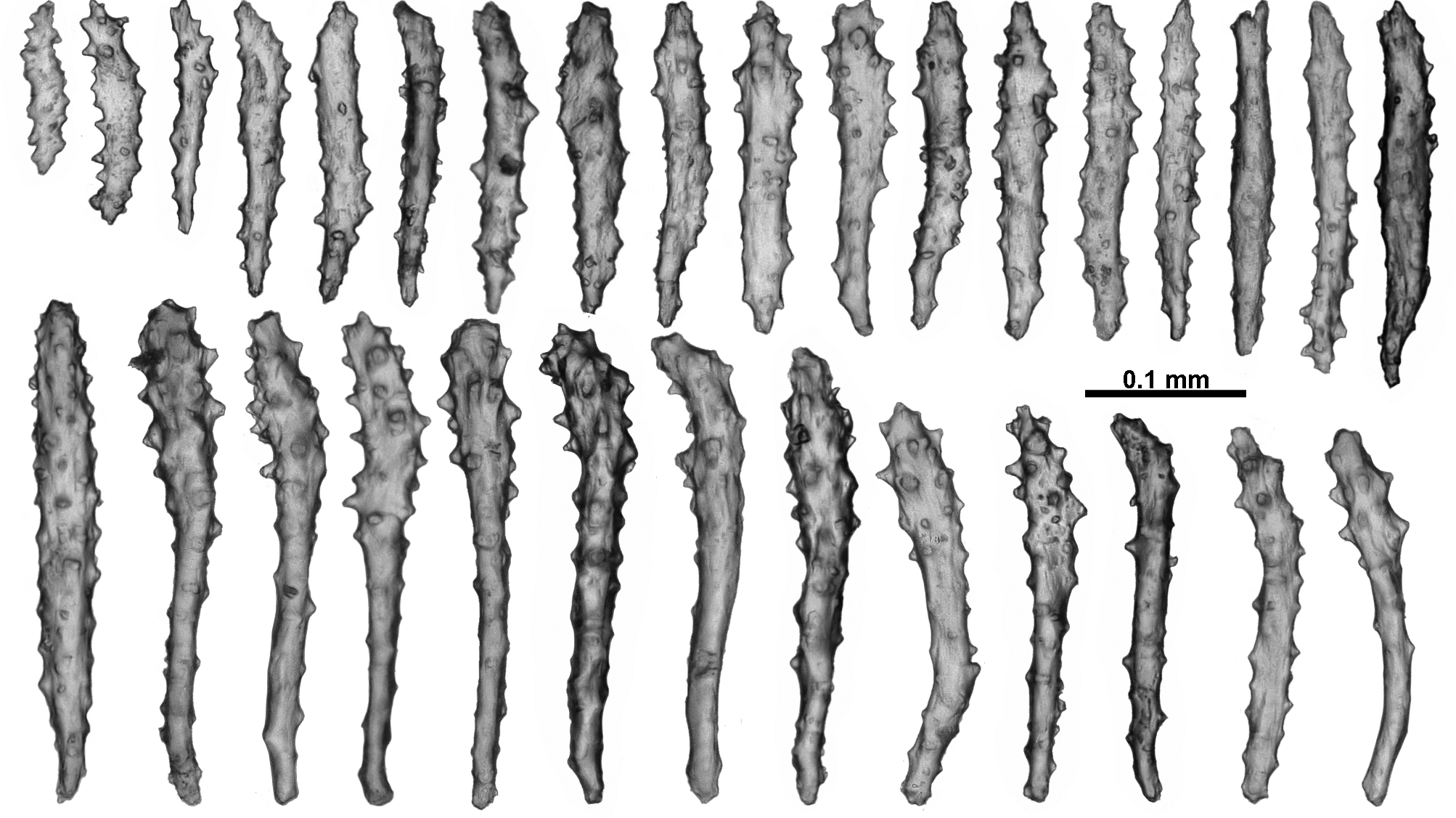
Sclerites: The polyp heads are well protected by being covered in crowded sclerites which are predominately long, warty sticks and spindles arranged as points and a bulky collaret. In the points, the sclerites are arranged en chevron, bunched so they are layered over each other ( Figs. 110 View FIGURE 110 C; 111A). They are mostly straight or slightly curved, narrow, sparsely tuberculate sticks and spindles, between 0.46–0.81 mm long ( Fig. 112 View FIGURE 112 A); tubercles are predominantly simple, blunt-topped or rounded cones. Among these are large, opaque, thick, prickly, club-shaped sclerites, approximately 0.45–0.77 mm long ( Fig. 113 View FIGURE 113 ). These occur more commonly in the distal part of the points and some of them continue longitudinally for a short distance along the tentacle rachis with the thickened blunt ends arranged distad ( Figs. 111 View FIGURE 111 A–C; 112B). They are usually straight although some bent spindles also occur ( Fig. 113 View FIGURE 113 a).
The collaret is composed of long curved spindles, arranged transversely in crowded bunches of between 10–20 on the widest part of the polyp head ( Fig. 111 View FIGURE 111 A), that have a comparable length to the more slender sclerites from the points. Below the collaret on the polyp neck are similar narrow, tuberculate spindles arranged mostly obliquely but more sparsely than in the collaret.
The tentacles fold over the mouth and have short, straight, thorny clubs and josephinae clubs crowded longitudinally along their aboral side ( Figs. 111 View FIGURE 111 B, C; 114). These grade in length from approximately 0.2–0.5 mm, with the length decreasing towards the tentacle tip, and with the clubbed ends positioned distad. Somewhat crowded, short spines cover the clubbed sclerite tip and some of the bent tips project up from the tentacle rachis giving it a rough surface.
Similarly, the pinnules have longitudinally arranged josephinae clubs, and the clubbed ends are directed towards the tip of the pinnules ( Fig. 112 View FIGURE 112 B). They are usually 0.25–0.35 mm long in the pinnules and possibly smaller than this in the rachis, although delineating in size between them is largely arbitrary ( Fig. 115 View FIGURE 115 ). Short and sparsely tuberculate sticks and spindles and flattened rods (0.11–0.24 mm long) are also crowded longitudinally in the pinnules.
No sclerites were found in the pharynx.
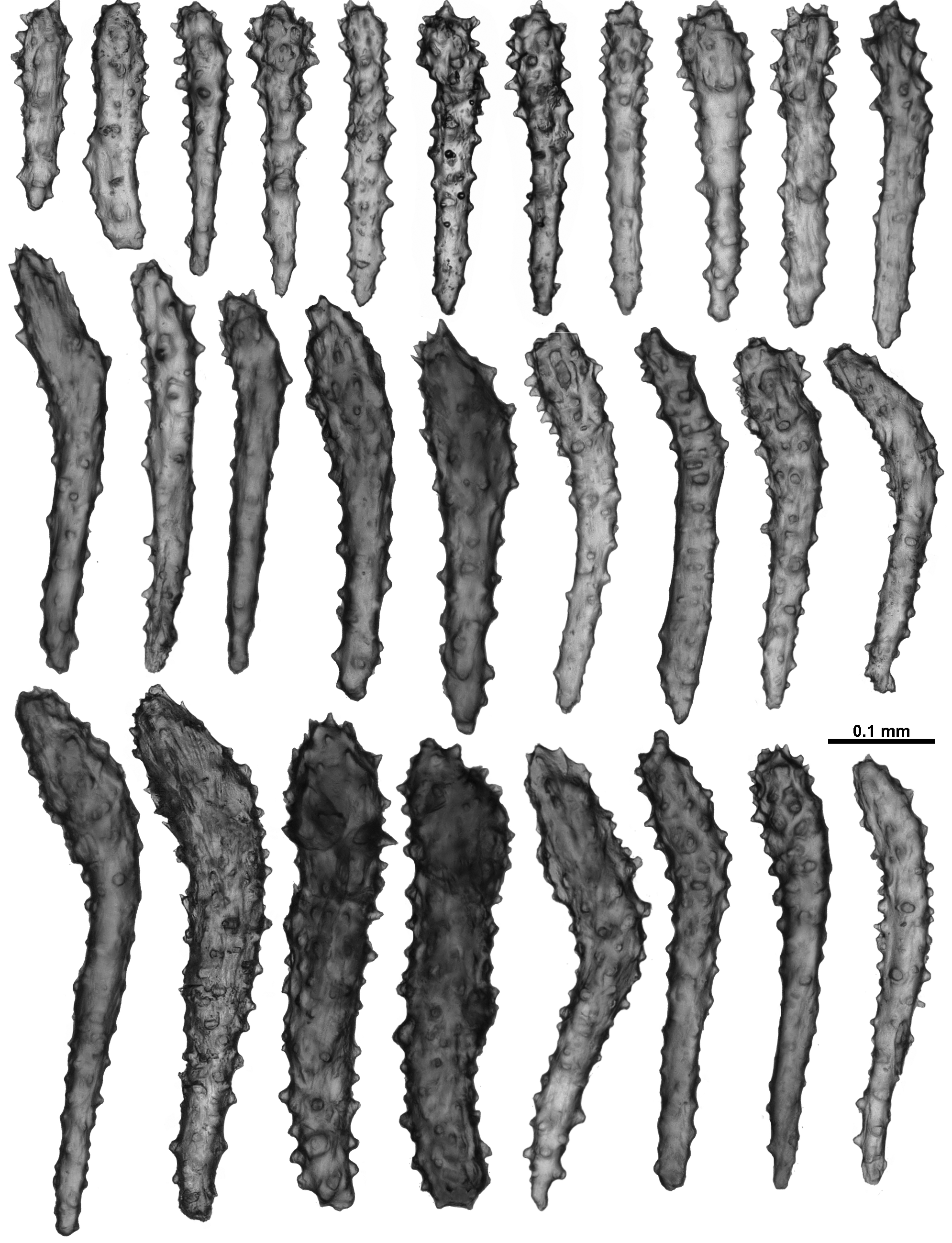
In the calyx, sclerites are in a single, almost translucent layer, and arranged transversely at the base, becoming en chevron distally such that there can be indistinct peaks of sclerites on the lip of the calyx ( Fig. 111 View FIGURE 111 A). The sclerites are long, mostly straight, sparsely tuberculate sticks and spindles, and are usually between 0.42–0.82 mm long ( Fig. 116 View FIGURE 116 ), although there are some shorter smoother spindles (0.27–0.37 mm).
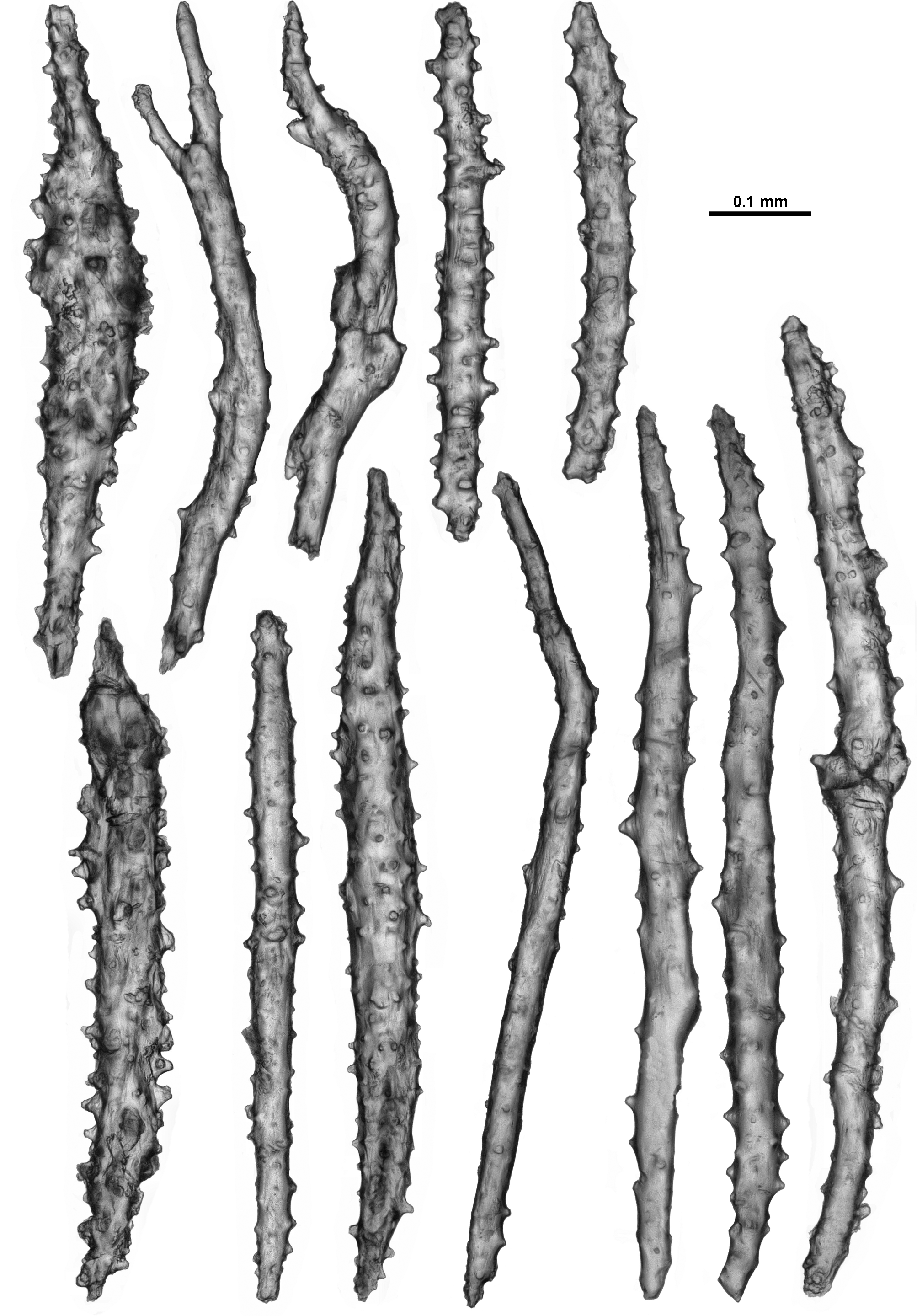
In the cortex, sclerites tend to be arranged longitudinally along the branch, although this arrangement is often interrupted or distorted by the calyces. They are quite crowded and form an opaque layer. Cortex sclerites are similar to the sclerites from the calyces—straight and curved sticks and spindles with simple tubercles, mixed with occasional small, smooth spindles ( Fig. 117 View FIGURE 117 ). They can be between 0.28–0.94 mm long but most are 0.43–0.86 mm.
The medulla is formed of tightly packed, longitudinally and obliquely placed sclerites that are most commonly long, straight or slightly bent sticks and spindles ( Fig. 118 View FIGURE 118 ). Amongst these are some sclerites with sparse tubercles, some with crowded, complex warts and thorns, and a few that are forked and branched, however no fused medulla sclerites were observed. The majority of the sclerites are between 0.47–0.98 mm but they can reach up to 1.25 mm.
Most sclerites are translucent under transmitted light except the thick clubs from the points and tentacles which are brown.
Variability: The only other specimen examined, MCZ 51046, has sclerites which are mostly similar to the holotype, but the colony form and the calyx distribution represent slight differences. The colony is in two pieces, each piece with a single bifurcation ( Fig. 119 View FIGURE 119 A). The largest consists of a branch portion, approximately 10 mm long, which then forks into two other branches, 21 mm and 35 mm long and 1.5–2.5 mm in diameter. The branches are slightly flattened in the plane of the colony and are oval in cross-section. Calyces are evenly and relatively sparsely distributed on both colony fragments, although they tend to form small clumps (maximum of 4 calyces) at the branch tip ( Fig. 119 View FIGURE 119 B). The calyces mostly emerge only on the narrow, lateral edges of the branches. When compared with the portions of the holotype of V. argentea n. comb., in MCZ 51046 the calyces are further apart, with much less clumping (the holotype has multiple crowded clumps of calyces) and the colony is finer. However, without knowing exactly how the complete holotype parent colony was constructed it is impossible to know the significance of these differences. The calyces have a single layer of sclerites arranged in indistinct chevrons, and the polyps have large clubbed sclerites arranged in the distal part of the points and on the tentacle rachis as in the holotype ( Fig. 119 View FIGURE 119 C, D). In addition, all other sclerites conform in general to those of the holotype, including the style of the josephinae clubs from the tentacles and pinnules ( Fig. 119 View FIGURE 119 E).
Distribution: The eastern Pacific Ocean (off the coast of Mexico) and the Hawaiian seamounts.
Depth: 1200–1559 metres.
Remarks: Despite the colony form and branching arrangement traditionally being of considerable taxonomic importance, Studer did not provide the colony size or an illustration, and the colony form cannot be reliably reconstructed from the pieces of the holotype. Nevertheless, Studer’s description of an “arborescente” branching arrangement with a main trunk supporting side branches is sufficiently different from the characteristic tangled colony form of Anthothela , with no main trunk and with common anastomoses, to be considered significant. Other features such as josephinae sclerites in the tentacles, obvious coelenteric canals in the medulla and the lack of sclerites in the pharynx mean this species cannot remain in Anthothela but does fit the diagnosis of Victorgorgia .
V. argentea n. comb. mostly resembles V. alba n. comb. (= A. nuttingi ) and V. macrocalyx n. comb., which also have large, warty clubs in the points. The most consistent difference is the sclerites from the tentacles—both those species have thick, warty rods as the main tentacle rachis sclerite and have poorly developed (or even lack) josephinae clubs, while V. argentea n. comb. has many josephinae clubs and lacks the warty rods.
The presence of large, opaque clubs in the distal part of the points and aboral side of the tentacles distinguishes this species from V. josephinae , V. nyahae n. sp. and V. eminens n. sp. Additionally, V. nyahae n. sp. has sharply pointed thorn clubs along the tentacle rachis and in the points.



| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Octocorallia |
|
Order |
|
|
SubOrder |
Scleraxonia |
|
Family |
|
|
Genus |
Victorgorgia argentea ( Studer, 1894 )
| Moore, Kirrily M., Alderslade, Philip & Miller, Karen J. 2017 |
Anthothela argentea
| Studer 1894: 60 |