
Uroplectes zambezicus, Prendini, 2015
|
publication ID |
https://doi.org/ 10.1206/3840.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5874503 |
|
persistent identifier |
https://treatment.plazi.org/id/03E5A832-FFBC-0537-A7C5-86BA34019753 |
|
treatment provided by |
Carolina |
|
scientific name |
Uroplectes zambezicus |
| status |
sp. nov. |
Uroplectes zambezicus View in CoL , sp. nov.
Figures 1 View FIGURE 1 , 2D, F View FIGURE 2 , 3C, D View FIGURE 3 , 4E, F View FIGURE 4 , 7B, D View FIGURE 7 , 8B, D View FIGURE 8 , 9B, D View FIGURE 9 , 12 View FIGURE 12 , 13 View FIGURE 13 ; table 2 View TABLE 2
Uroplectes chubbi Hirst, 1911 View in CoL : FitzPatrick, 1996: 64; FitzPatrick, 2001: 191 [misidentification: NHMZ S84/89, S85/17, S86/5, S86/71, S90/17, S95/129].
HOLOTYPE: MOZAMBIQUE: Tete Province: Tete District: Zambezi River, N side, 2.4 km towards Chiuta on road EN222 from junction with road EN103 (Tete – Zòbué ), 16°06′50″S 33°37′01″E, 150 m, 10.xii.2007, L. Prendini and W. R. Schmidt, arid mopane savanna on alluvial silty-loam soil with scattered shale and sandstone outcrops, taken under small stone during day, 1 ♂ ( AMNH). GoogleMaps
PARATYPES: MOZAMBIQUE: Tete Province: Moatize District: Tenge Hill , 15°43.424′S 33°46.313′E, 361 m, 22–24.iii.2013, P.G. Hawkes and J.N. Fisher, 2 ♀ ( AMNH), 6.ix.2013, P.G. Hawkes and J.N. Fisher, day collecting, 11 ♂, 1 ♀ ( AMNH), night collecting with UV, 18:30– 20:30, 1 ♂ ( AMNH), 7.ix.2013, P.G. Hawkes and J.N. Fisher, day collecting, 1 ♂, 2 ♀ ( AMNH) GoogleMaps ; 15°44.900′S 33°46.159′E, 237 m, 7.ix.2013, P.G. Hawkes and J.N. Fisher, mopane woodland, night collecting with UV, 19:00, 1 ♀ ( AMNH). GoogleMaps Tete District: Zambezi River , N side, 2.4 km towards Chiuta on road EN222 from junction with road EN103 ( Tete – Zòbué ), 16°06′50″S 33°37′01″E, 150 m, 10.xii.2007, L. Prendini and W. R. Schmidt , arid mopane savanna on alluvial silty-loam soil with scattered shale and sandstone outcrops, collected with UV detection on warm and humid, still, dark night, running or walking on surface, mainly in leaf-litter around base of bushes and in rocky areas, syntopic with Hottentotta trilineatus ( Peters, 1861) , 11 ♂, 7 ♀ ( AMNH), 3 first instars ( AMCC [ LP 8230]); Tete, base of hill with communications tower overlooking town (at bend in road), 16°12′58.9″S 33°34′27.7″E, 189 m, 10.xii.2007, L. Prendini and W. R. Schmidt, arid savanna with baobabs on alluvial silty-loam soil, fairly dense vegetation cover, UV light detection on warm and humid, dark night, running on ground surface, especially in leaf-litter under tree canopy, syntopic with H. trilineatus and Uroplectes planimanus ( Karsch, 1879) , 4 ♂, 3 ♀ ( AMNH), 1 ♂ ( AMCC [ LP 8229]); Tete, midslope on hill with communications tower overlooking town, 16°12′55″S 33°34′12″E, 245 m, 10.xii.2007, L. Prendini and W. R. Schmidt, arid savanna with baobabs, UV detection on warm and humid, dark night, on rocky slopes along road, sympatric with Hadogenes troglodytes (Peters, 1861) , H. trilineatus and U. planimanus , 1 ♂ ( AMNH). ZAMBIA: Lusaka Province: Chiawa, junction of Kafue and Zambezi rivers, 15°53.011′S 28°53.942′E, 442 m, 21.iv.2015, W. Conradie, mopane woodland, caught in reptile trap (bucket pitfalls), 1 ♂, 1 ♀ ( AMNH), 1 ♂ ( AM). ZIMBABWE: Mashonaland Central Province: Guruve District: Guruve, 4 km NE Gonono School [16°05′S 30°50′E], 28.iii.1997, F. Nyathi, under log, 1 ♀ ( NHMZ S97/21), 31.iii.1997, F. Nyathi, pitfall traps, 1 ♂ ( NHMZ S97/2); Guruve, 7 km E Mushumbi Pools [16°06′S 30°32′E], 22.iii.1997, F. Nyathi, under mopane bark, 3 ♀ ( NHMZ S97/11). Mashonaland West Province: Hurungwe District: Hurungwe Safari Area: Rifa Conservation Camp [16°10′S 28°50′E], 4–9.xii.1995, Girls’ College and NHMZ, pitfall traps, 10 ♂, 4 ♀, 1 subad. ♀, 1 juv. ♂, 2 first instars ( NHMZ S 96/9). Mana Pools National Park: Nyakasikana Fly Gate, Mana Pools [16°03′S 29°24′E], 9.xii.1984, Falcon College Expedition, 1 ♀ ( NHMZ S 86/5). Kariba District: Matusadona National Park: Chin- gachereyari, Matusadona, 16°44′S 28°40′E, 13.viii.1983, Zimbabwe Schools Expedition Society, 1 subad. ♂ ( NHMZ S 83/141); Mbizi Pan, Matusadona [16°49′S 28°28′E], 12.xii.1983, G. Putterill, 1 ♂ ( NHMZ S 84/89). Matabeleland North Province: Hwange District: Chisuma area [18°01′S 25°57′E], 31.vii.1990, F. Nyathi, 2 ♂, 1 subad. ♂, 2 subad. ♀ ( NHMZ S 90/17); Dete [18°38′S 26°52′E], 6.ii.1985, D. Adams, 1 ♀ ( NHMZ S 85/17); Mzola Camp [18°30′S 27°24′E], 4.xii.1997, Girls’ College and NHMZ, 1 ♀ ( NHMZ S 13/4 ex S 99/20); Mzola Camp, 2 km N Homestead [18°30′S 27°24′E], 11.xii.1997, Girls’ College and NHMZ, under bark, 2 ♀ ( NHMZ S 99/16, S 99/17). Hwange National Park: Robin’s Camp [18°38′S 25°59′E], 1.ii.1986, N. English, 1 ♀ ( NHMZ S 86/71) ; Wankie [Hwange], 18°22′S 26°29′E, v.1961, D.G. Broadley, 1 ♀ ( NMSA 8319). Matetsi Safari Area: Kasetsheti Weirs, Matetsi [18°18′S 25°48′E], 11.x.1988, F. Nyathi, 1 ♂ ( NHMZ S 89/3); Rosslyn Camp, Matetsi, Tshowe River [18°30′S 25°55′E], 5–10.xii.1994, Girls’ College and NHMZ, 1 ♀ ( NHMZ S 95/129).
ETYMOLOGY: The specific epithet is derived from the Zambezi River Valley, where all the known localities of the new species are located.
DIAGNOSIS: Uroplectes zambezicus , sp. nov., appears to be most closely related to U. malawicus , sp. nov., based on the lower pectinal tooth count (15–17) and enlarged basal pectinal tooth of the female in both species, by means of which they may be separated from U. katangensis , sp. nov., in which the pectinal tooth count is higher (18–19) and the basal pectinal tooth unmodified in the female. Uroplectes zambezicus , sp. nov., differs from U. malawicus , sp. nov., and U. katangensis , sp. nov., in possessing a slightly broader chela manus and a shorter, broader metasoma (more so in the adult male) as well as less uniform coloration. The tergites exhibit a narrow immaculate stripe medially and sometimes a pair of narrow immaculate stripes laterally in U. zambezicus , sp. nov., but are almost entirely infuscate in U. malawicus , sp. nov., and U. katangensis , sp. nov. The chelicerae, pedipalp trochanter, femur, patella, and legs are entirely immaculate in U. zambezicus , sp. nov., whereas the dorsal surfaces of the chelicerae, lateral surfaces of the pedipalp patella, and often dorsal and prodorsal surfaces of the pedipalp femur and trochanter, as well as dorsal and retrolateral surfaces of the leg femora, patellae, and tibia are partially infuscate in U. malawicus , sp. nov. Metasomal segment IV is often paler in U. zambezicus , sp. nov., than in U. malawicus , sp. nov. Uroplectes zambezicus , sp. nov., differs further from U. katangensis , sp. nov., in the lower counts of median denticle subrows on the fixed and movable fingers of the pedipalp chela (11 or 12 vs. 13, respectively), and from U. malawicus , sp. nov., in the less numerous and pronounced spiniform granules in the distal half of the chela manus prodorsal surface in the adult male.
DESCRIPTION: The following account, which is based on the type material, describes only those characters that differ from U. malawicus , sp. nov.
Total length: Adult: small, maximum length, measured from anterior margin of carapace to tip of aculeus, 23 mm (20–27 mm, n = 6) (♂), 26 mm (24–27 mm, n = 6) (♀) ( table 2 View TABLE 2 ).
Color: As for U. malawicus , sp. nov., except as follows (figs. 2D, F, 12). Chelicerae, pedipalp trochanter, femur, patella, and legs entirely immaculate. Carapace mostly infuscate except for a pair of narrow immaculate stripes laterally in some specimens. Tergites mostly infuscate except for narrow immaculate stripe medially and, in some specimens, a pair of narrow immaculate stripes laterally Carapace: As for U. malawicus , sp. nov., except as follows (fig. 3C, D). Anterior width of posterior width, 52% (51%–46%, n = 6) (♂), 56% (47%–78%, n = 6) (♀) ( table 2 View TABLE 2 ); posterior width of length, 104% (96%–110%, n = 6) (♂), 106% (75%–119%, n = 6) (♀). Median ocular tubercle, distance from anterior carapace margin 41% (n = 6) (♂), 40% (n = 6) (♀) of carapace length.
.
Pedipalps: As for U. malawicus , sp. nov., except as follows (fig. 13). Femur width of length, 32% (30%–34%, n = 6) (♂), 34% (30%–36%, n = 6) (♀) ( table 2 View TABLE 2 ). Patella width of length, 37% (35%–40%, n = 6) (♂), 39% (38%–40%, n = 6) (♀). Chela manus, width of height, 102% (100%–107%, n = 6) (♂), 108% (100%–116%, n = 6) (♀); width of length along ventroexternal carina, 57% (54%–62%, n = 6) (♂), 62% (56%–67%, n = 6) (♀); length along ventroexternal carina of length movable finger, 68% (59%–90%, n = 6) (♂), 57% (53%–63%, n = 6) (♀). Manus surfaces smooth except prodorsal surface with relatively dense spiniform granules, more numerous and pronounced in distal half (♂) (fig. 13E–H) or few small scattered spiniform granules (♀). Fixed and movable fingers, median denticle rows respectively comprising 11 (11–12, n = 12) and 12 (12–13, n = 12) oblique denticle subrows.
Pectines: As for U. malawicus , sp. nov., except as follows (fig. 4E, F). Pectinal teeth curved, all similar in size (♂) or basal pectinal tooth enlarged, approximately twice the size of other teeth (♀); tooth count, 16/17 (15–17/15–18, n = 12) (♂), 16/16 (15–17/16–17, n = 12) (♀) ( table 2 View TABLE 2 ).
Mesosoma: As for U. malawicus , sp. nov., except as follows. Sternite VII, length of width, 62% (49–74%, n = 6) (♂), 53% (48–57%, n = 6) (♀) ( table 2 View TABLE 2 ).
Metasoma and telson: As for U. malawicus , sp. nov., except as follows (figs. 7B, D, 8B, D, 9B, D). Metasomal segment V, width of segment I, width, 109% (107%–114%, n = 6) (♂), 109% (105%–113%, n = 6) (♀) ( table 2 View TABLE 2 ). Metasoma width of length, segment I, 92% (84%– 98%, n = 6) (♂), 107% (103%–110%, n = 6) (♀); II, 81% (78%–88%, n = 6) (♂), 96% (93%–102%, n = 6) (♀); III, 80% (75%–88%, n = 6) (♂), 92% (87%–96%, n = 6) (♀); IV, 74% (70%–83%, n = 6) (♂), 84% (80%–87%, n = 6) (♀); V, 71% (67%–79%, n = 6) (♂), 81% (80%–85%, n = 6) (♀). Telson vesicle, width of metasomal segment V, 62% (57%–66%, n = 6) (♂), 64% (58%–86%, n = 6) (♀); height of length, 64% (60%–68%, n = 6) (♂), 65% (62%–70%, n = 6) (♀). Aculeus length of vesicle length, 70% (65%–76%, n = 6) (♂), 69% (59%–81%, n = 6) (♀). Length metasoma and telson, of total length, 59% (56%–60%, n = 6) (♂), 56% (55%–57%, n = 6) (♀).
DISTRIBUTION: Presently recorded from five localities in Mozambique, one in Zambia and thirteen in Zimbabwe (fig. 1). All known locality records fall within the Zambezi River Valley.
ECOLOGY: The known material was collected in arid savanna, dominated by the mopane tree, Colophospermum mopane (Benth.) J. Léonard , in the Zambezi River Valley. Zimbabwean material deposited in the NHMZ was collected under tree bark and logs, and in pitfall traps. Recently collected material originates from three localities at elevations of 150–360 m in the vicinity of Tete, Mozambique, and a fourth locality, at an elevation of 442 m, north of the junction of the Kafue and Zambezi rivers, in Zambia. The two Mozambican localities at which material was collected by the author differ in habitat. The habitat near Tete was fairly dense mixed broadleaf woodland with baobab trees, Adansonia digitata L. (fig. 2C), on alluvial silty-loam soil, around the base and on the lower slopes of a rocky hill. Specimens were collected with UV light detection on a warm and humid, moonless night, running on the ground surface, especially in leaf litter under the tree canopy (fig. 2E). The habitat on the north side of the Zambezi River was more open, arid mopane woodland with scattered shale and sandstone outcrops, also on alluvial silty-loam soil. One specimen was found under a small stone during the day. The others were found with UV detection on a warm, still, moonless, humid night, running or walking on the ground surface, mainly in leaf litter around the base of bushes and in rocky areas. The material from Zambia was collected in reptile traps (bucket pitfall traps) in mopane woodland (W. Conradie, personal commun.). The habitat and habitus (figs. 2D, F, 12) of U. zambezicus , sp. nov., are consistent with the lapidicolous and corticolous ecomorphotypes ( Prendini, 2001). Five buthids, Hottentotta trilineatus , Parabuthus mossambicensis (Peters, 1861) , Uroplectes flavoviridis , U. planimanus , and U. xanthogrammus , a hormurid, Hadogenes troglodytes , and two scorpionids, Opistophthalmus carinatus (Peters, 1861) and O. glabrifrons , were collected in sympatry at one or more of the collection localities in Mozambique and Zimbabwe.
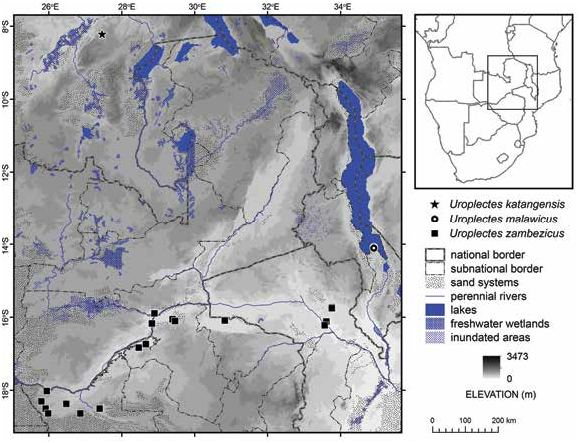

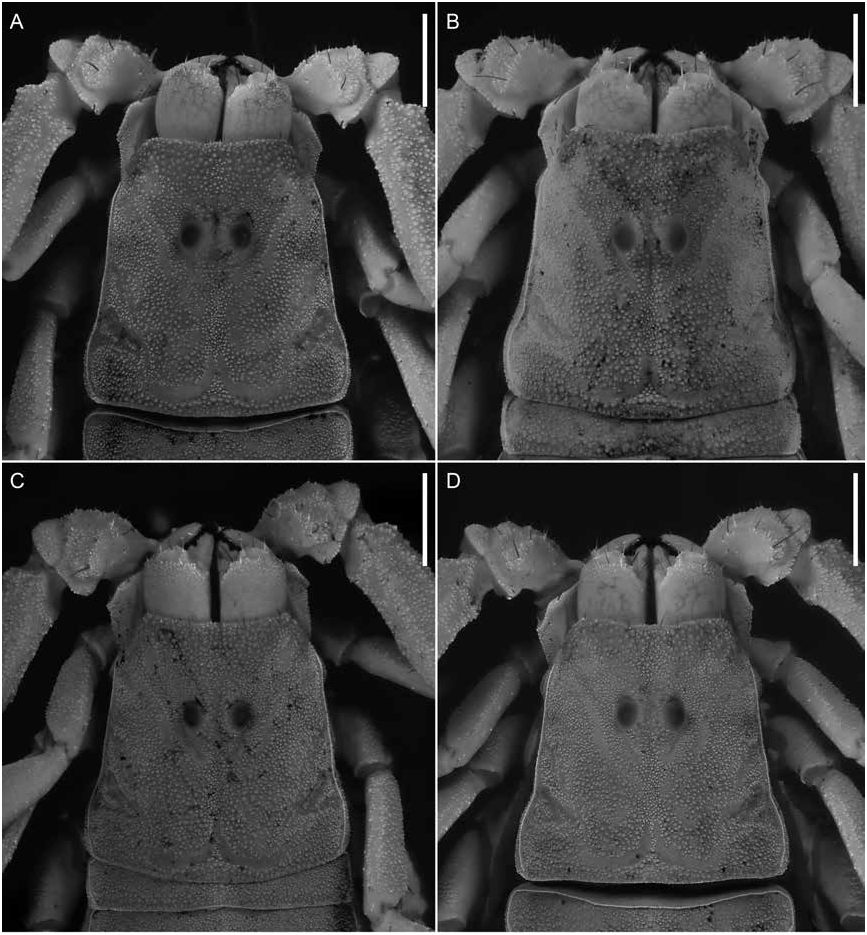
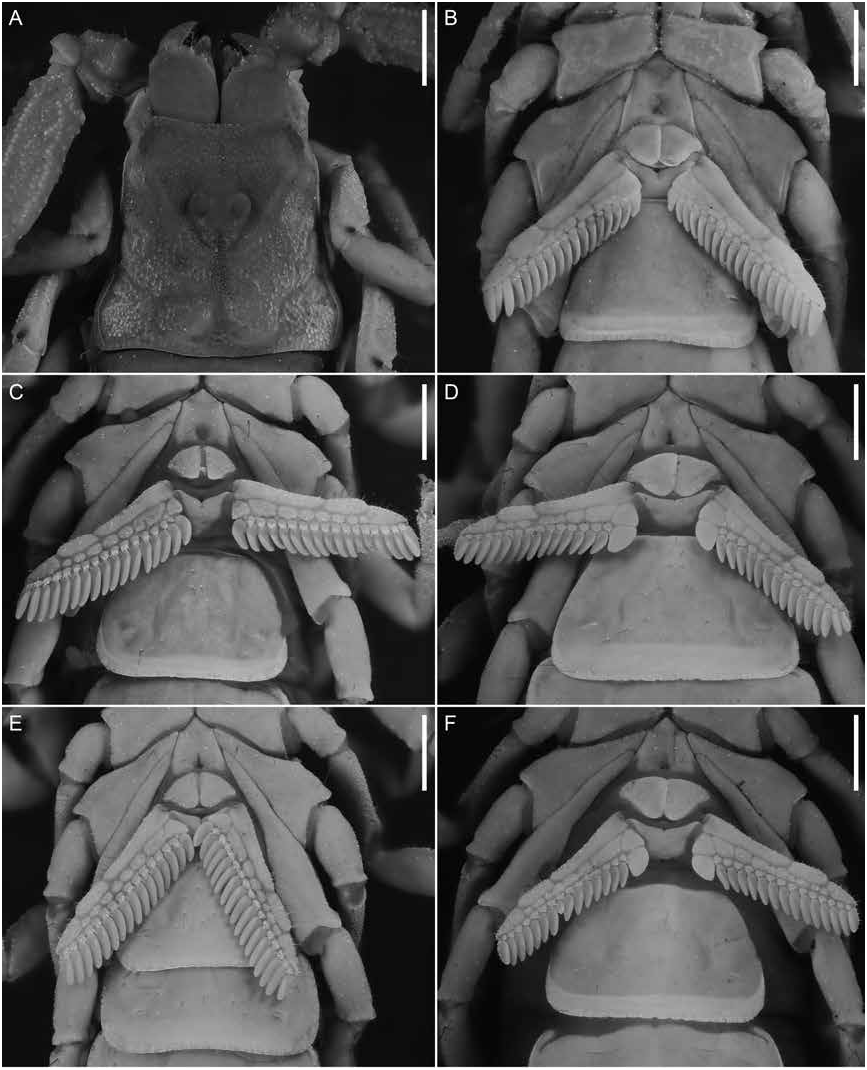
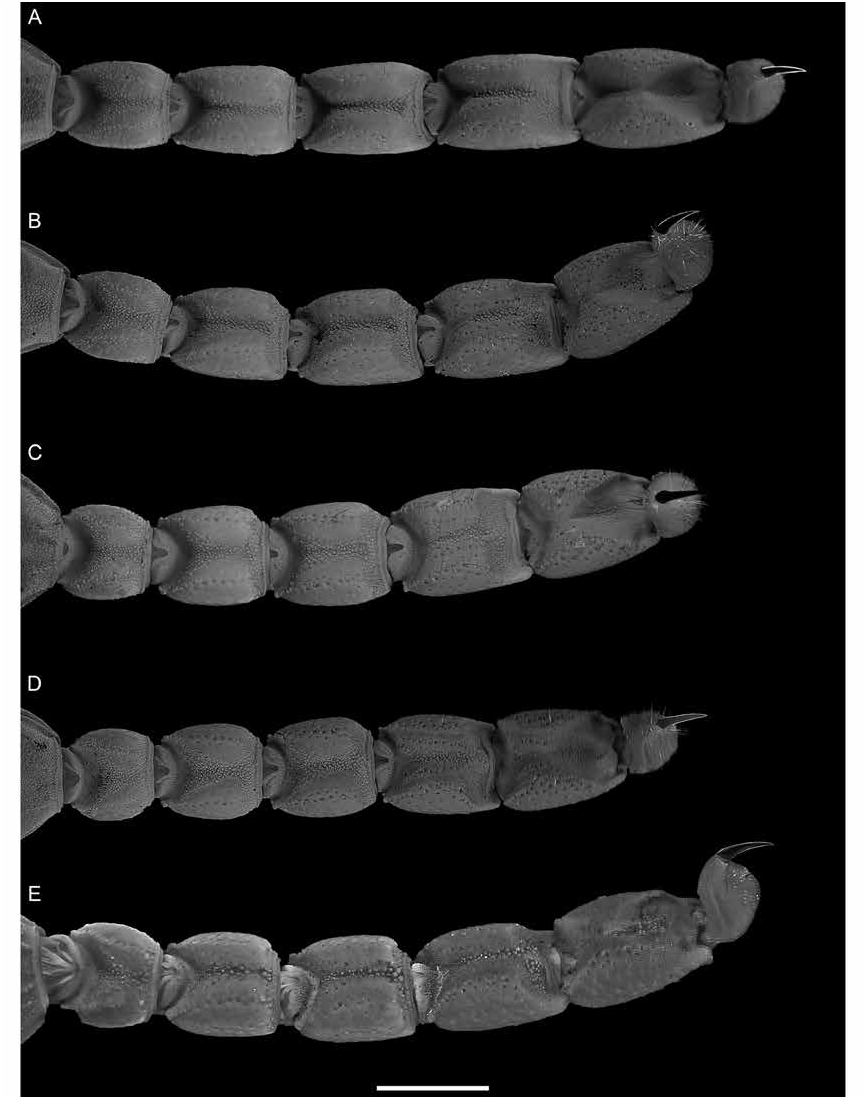


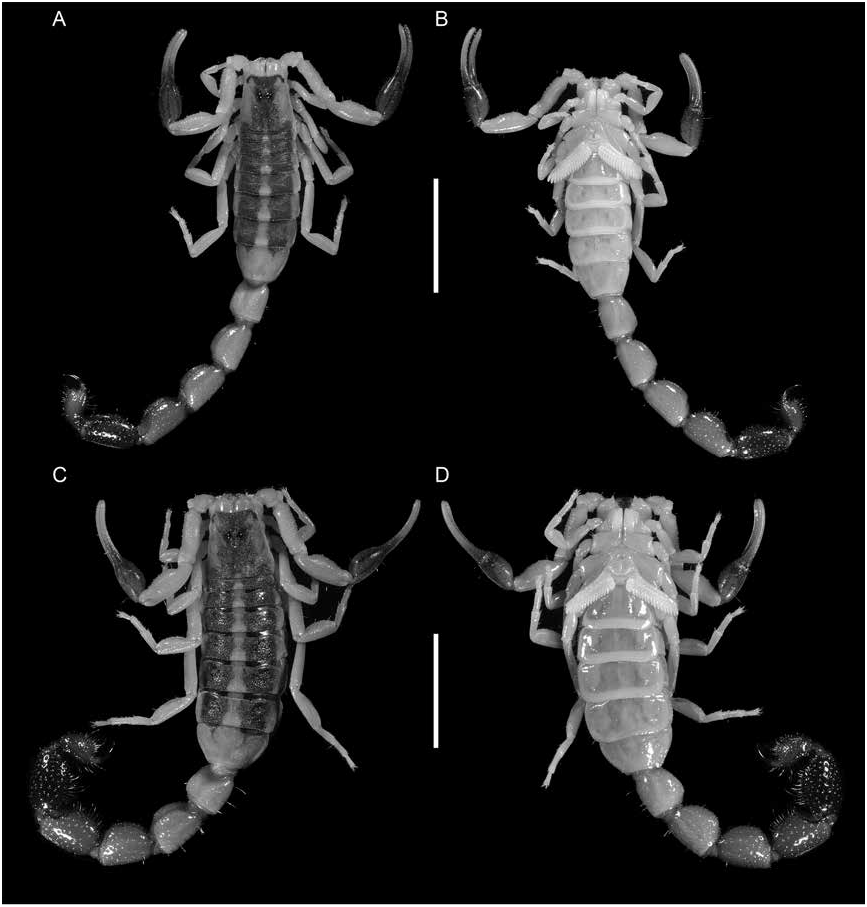
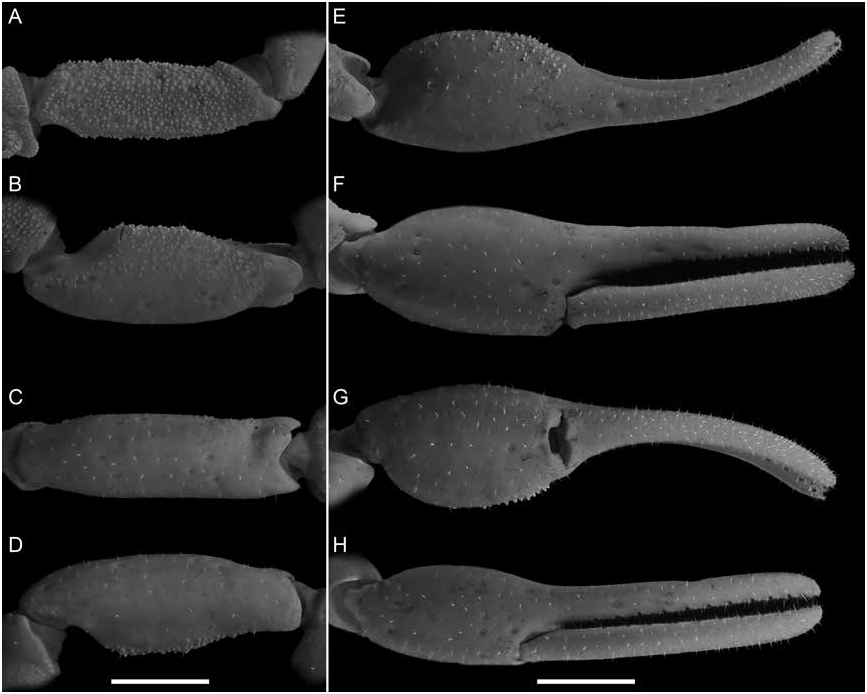
TABLE 2. Meristic data for six males and six females of Uroplectes zambezicus, sp. nov., in the collections of the American Museum of Natural History, New York (AMNH) and the Natural History Museum of Zimbabwe, Bulawayo (NHMZ). Measurements (mm) follow Prendini (2004a, 2004b, 2004c, 2005b). 1Sum of carapace, tergites I–VII, metasomal segments I–V, and telson; 2distance from anterior carapace margin; 3sum of metasomal segments I–V and telson; 4distance from base of condyle to tip of fixed finger.
| Specimen | sex | ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♀ | ♀ | ♀ | ♀ | ♀ | ♀ |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| type | Holo. | Para. | Para. | Para. | Para. | Para. | Para. | Para. | Para. | Para. | Para. | Para. | |
| coll. | AMNH | AMNH | NHMZ | NHMZ | NHMZ | NHMZ | AMNH | AMNH | NHMZ | NHMZ | NHMZ | NHMZ | |
| number | S97/2 | S84/89 | S90/17 | S89/3 | S99/17 | S86/5 | S85/17 | S97/11 | |||||
| Total length 1 | 22.6 | 26.0 | 22.0 | 20.8 | 26.6 | 20.2 | 25.5 | 26.0 | 24.6 | 27.2 | 24.4 | 26.8 | |
| Carapace | median ocelli 2 | 0.9 | 1.1 | 0.8 | 0.7 | 1.0 | 0.8 | 1.0 | 1.0 | 0.9 | 1.0 | 0.9 | 1.1 |
| length | 2.5 | 3.0 | 2.4 | 2.1 | 2.9 | 2.2 | 2.8 | 2.8 | 2.7 | 3.0 | 2.6 | 2.8 | |
| anterior width | 1.3 | 1.7 | 1.3 | 1.2 | 1.5 | 1.2 | 1.7 | 1.7 | 1.6 | 1.7 | 1.5 | 1.7 | |
| posterior width | 2.5 | 3.3 | 2.5 | 2.2 | 2.8 | 2.3 | 3.1 | 3.1 | 2.0 | 3.3 | 3.1 | 3.1 | |
| Tergite I | length | 0.4 | 0.6 | 0.4 | 0.5 | 0.5 | 0.4 | 0.6 | 0.6 | 0.5 | 0.5 | 0.5 | 0.5 |
| Tergite II | length | 0.6 | 0.8 | 0.6 | 0.7 | 0.7 | 0.6 | 0.8 | 0.8 | 0.8 | 0.9 | 0.8 | 0.8 |
| Tergite III | length | 0.8 | 1.0 | 0.8 | 0.9 | 1.0 | 0.8 | 1.2 | 1.2 | 1.1 | 1.2 | 1.1 | 1.1 |
| Tergite IV | length | 1.0 | 1.2 | 1.0 | 1.0 | 1.2 | 0.9 | 1.3 | 1.4 | 1.3 | 1.4 | 1.3 | 1.4 |
| Tergite V | length | 1.1 | 1.3 | 1.1 | 1.1 | 1.4 | 1.0 | 1.4 | 1.5 | 1.4 | 1.5 | 1.5 | 1.6 |
| Tergite VI | length | 1.2 | 1.4 | 1.2 | 1.2 | 1.5 | 1.1 | 1.5 | 1.6 | 1.5 | 1.6 | 1.6 | 1.6 |
| Tergite VII | length | 1.7 | 1.6 | 1.6 | 1.5 | 2.0 | 1.5 | 1.8 | 1.8 | 1.9 | 2.0 | 1.8 | 1.8 |
| Sternite VII | length | 1.6 | 1.6 | 1.4 | 1.4 | 2.0 | 1.2 | 1.7 | 1.6 | 1.7 | 1.8 | 1.4 | 1.8 |
| width | 2.4 | 3.3 | 2.2 | 2.2 | 2.7 | 2.0 | 3.2 | 3.1 | 3.0 | 3.3 | 2.9 | 3.2 | |
| Mesosoma | total length | 6.7 | 7.8 | 6.5 | 6.7 | 8.3 | 6.3 | 8.5 | 8.8 | 8.3 | 9.1 | 8.4 | 8.7 |
| Metasoma I | length | 1.8 | 2.1 | 1.9 | 1.6 | 2.1 | 1.5 | 1.9 | 1.9 | 1.8 | 2.0 | 1.8 | 2.0 |
| width | 1.6 | 2.0 | 1.6 | 1.5 | 1.9 | 1.4 | 2.0 | 2.0 | 1.9 | 2.2 | 1.9 | 2.1 | |
| height | 1.3 | 1.6 | 1.3 | 1.3 | 1.5 | 1.2 | 1.6 | 1.6 | 1.6 | 1.7 | 1.6 | 1.7 | |
| Metasoma II | length | 2.1 | 2.4 | 2.1 | 1.9 | 2.5 | 1.9 | 2.1 | 2.2 | 2.1 | 2.4 | 2.0 | 2.3 |
| width | 1.7 | 2.1 | 1.7 | 1.5 | 2.0 | 1.5 | 2.2 | 2.1 | 2.0 | 2.3 | 1.9 | 2.2 | |
| height | 1.4 | 1.7 | 1.4 | 1.4 | 1.6 | 1.3 | 1.8 | 1.7 | 1.6 | 1.8 | 1.6 | 1.7 | |
| Metasoma III | length | 2.1 | 2.5 | 2.2 | 2.0 | 2.6 | 2.0 | 2.4 | 2.4 | 2.3 | 2.5 | 2.1 | 2.6 |
| width | 1.8 | 2.2 | 1.8 | 1.6 | 2.0 | 1.5 | 2.3 | 2.2 | 2.1 | 2.4 | 2.0 | 2.3 | |
| height | 1.5 | 1.7 | 1.5 | 1.5 | 1.7 | 1.4 | 1.9 | 1.8 | 1.6 | 1.8 | 1.6 | 1.8 |
| NMSA |
South Africa, Kwa-Zulu Natal, Pietermaritzburg, Natal Museum |
| R |
Departamento de Geologia, Universidad de Chile |
| AMNH |
American Museum of Natural History |
| UV |
Departamento de Biologia de la Universidad del Valle |
| AMCC |
Ambrose Monell Cryo Collection, American Museum of Natural History |
| LP |
Laboratory of Palaeontology |
| AM |
Australian Museum |
| NMSA |
KwaZulu-Natal Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Uroplectes zambezicus
| Prendini, Lorenzo 2015 |
Uroplectes chubbi
| Hirst 1911 |