
Trucidocynodon riograndensis Oliveira, Soares & Schultz, 2010
|
publication ID |
https://doi.org/10.11646/zootaxa.4457.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:3115F43D-B088-4E4C-9385-51FF04C4C7D9 |
|
DOI |
https://doi.org/10.5281/zenodo.5980189 |
|
persistent identifier |
https://treatment.plazi.org/id/807DD96E-841F-6A74-FF21-FA0070D7FBC2 |
|
treatment provided by |
Plazi |
|
scientific name |
Trucidocynodon riograndensis Oliveira, Soares & Schultz, 2010 |
| status |
|
Trucidocynodon riograndensis Oliveira, Soares & Schultz, 2010
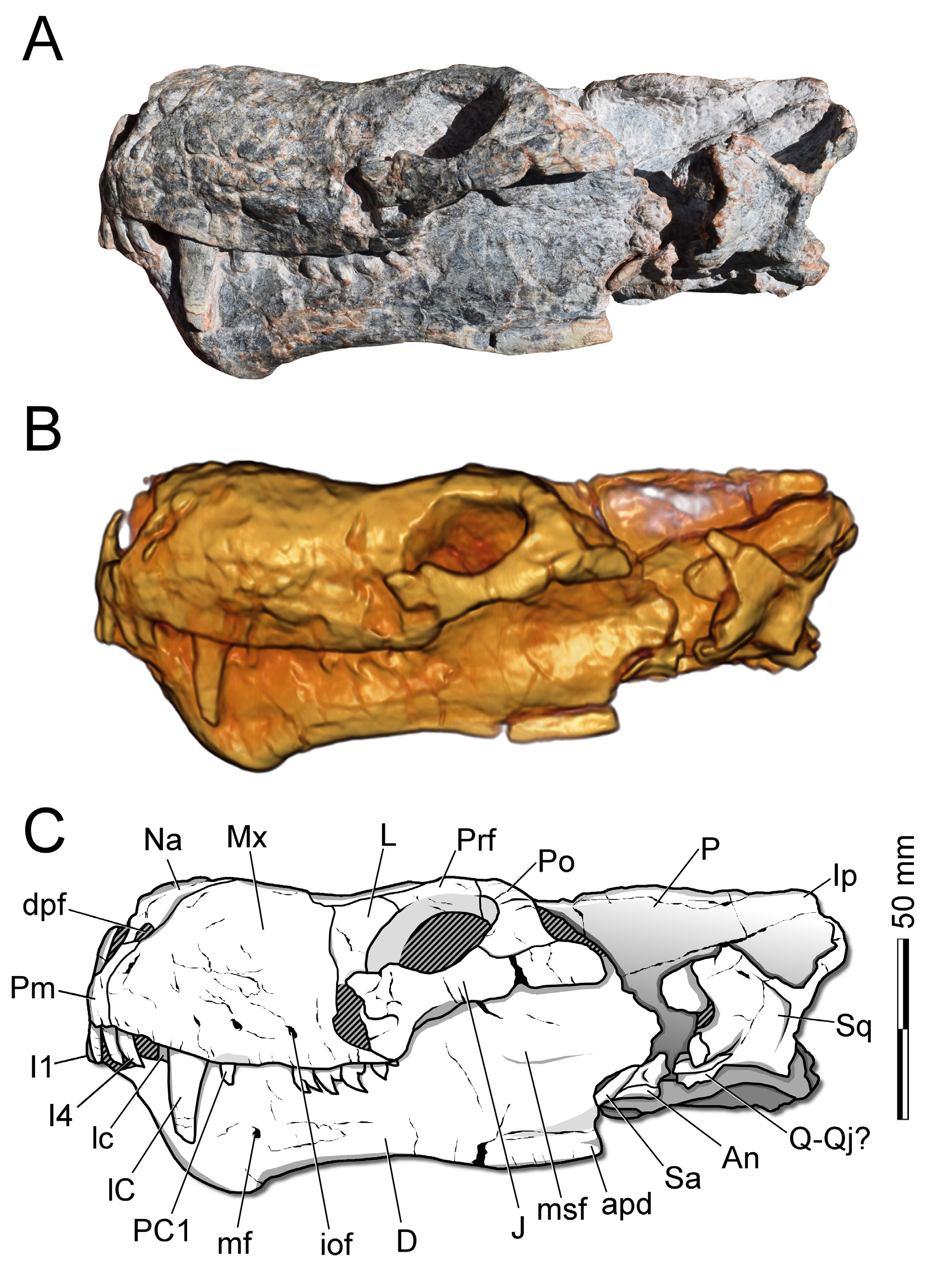
( FIGS. 2–8 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 ; TAbLE 1)
Holotype: UFRGS PV-1051-T, ALMOST COMPLETE SkELETON (OLIVEIRA et al. 2010).
Paratypes: UFRGS PV-1053-T, RIGHT SCAPULAR GIRDLE, AND fORELIMb; UFRGS PV-1069-T, fIVE VERTEbRAE, AND SOME fRAGMENTARY RIbS; UFRGS PV-1070-T, A RIGHT TIbIA; UFRGS PV-1071-T, RIGHT ULNA (OLIVEIRA et al. 2010).
Diagnosis: SEE OLIVEIRA et al. 2010.
New Referred specimen: CAPPA /UFSM 0 0 29, A COMPLETE SkULL INCLUDING ARTICULATED LOWER jAW.
Locality and horizon: CAPPA /UFSM 0 0 29 COMES fROM THE JANNER OUTCROP, MUNICIPALITY Of AGUDO, STATE Of RIO GRANDE DO SUL, BRAzIL, WHICH IS INCLUDED IN THE LOWER PORTION Of THE CANDELáRIA SEqUENCE (HORN et al. 2014), Hyperodapedon AZ, SANTA MARIA SUPERSEqUENCE (ZERfASS et al. 2003).
Description and comparison:
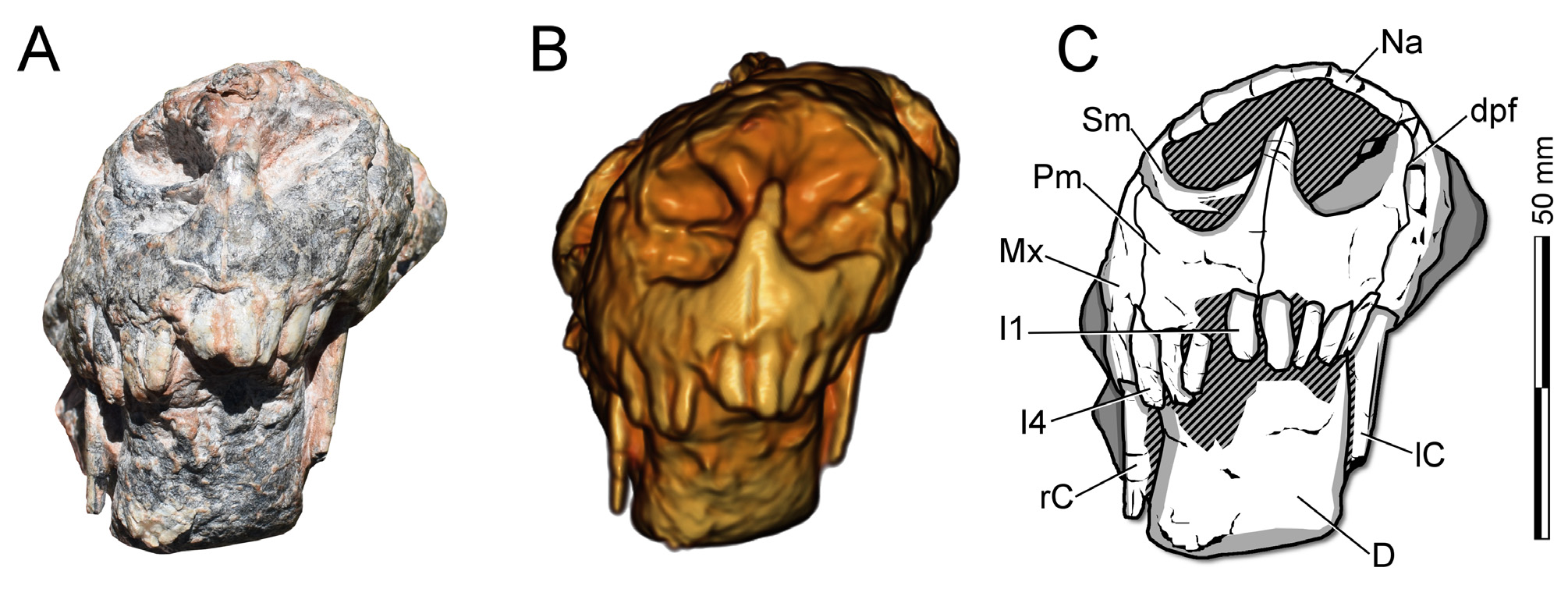
Premaxilla. CAPPA / UFSM 0 0 29 PRESERVES bOTH PREMAXILLAE ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ) . THE MAIN bODY Of THIS ELEMENT IS PROPORTIONALLY LARGER THAN IN TRAVERSODONTID CYNODONTS (E.G., Exaeretodon riograndensis ), WITH PROPORTIONS RESEMbLING THOSE Of OTHER ECTENINIIDS. THE INTERNARIAL PROCESS Of CAPPA/UFSM 0 0 29 IS TALL bUT DOES NOT REACH THE NASALS ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). IN LATERAL VIEW, IT IS DORSALLY DIRECTED IN ITS VENTRAL HALf, WHEREAS THE DORSAL PART IS DORSOPOSTERIORLY DIRECTED, SIMILARLY TO Ecteninion lunensis . THIS STRUCTURE IS SIGMOID IN LATERAL VIEW IN Diegocanis
elegans (ALTHOUGH THIS COULD bE TAPHONOMIC), AND IN THE HOLOTYPE Of Trucidocynodon riograndensis IT IS MISSING (OLIVEIRA et al. 2010). THE POSTEROLATERAL ASCENDING PROCESS IS LONG AND REACHES THE DORSAL LEVEL Of THE EXTERNAL NARES, bUT DOES NOT CONTACT THE NASAL, SIMILAR TO THE CONDITION Of HOLOTYPE UFRGS PV-1051-T AND Chiniquodon sanjuanensis (PVSJ 411). IN CONTRAST, IT IS SHORT IN E. lunensis AND D. elegans (MARTÍNEz et al. 2013). THE POSTEROVENTRAL bORDER Of THE PREMAXILLA CONTACTS THE ANTEROVENTRAL MARGIN Of THE MAXILLA AT THE LEVEL Of THE fOURTH UPPER INCISOR, AS IN THE HOLOTYPE Of T. riograndensis , E. lunensis , AND D. elegans (MARTÍNEz et al. 1996; 2013; OLIVEIRA et al. 2010). IN DORSAL VIEW, THE POSTEROLATERAL ASCENDING PROCESS Of THE PREMAXILLA Of CAPPA/UFSM 0 0 29 ENDS AT THE ANTERIOR bORDER Of THE PARACANINE fOSSA, WHICH IS OPENED DORSALLY ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 , 8 View FIGURE 8 ).
Septomaxilla. BOTH SEPTOMAXILLAE ARE ALMOST COMPLETE, AND ONLY THE LEfT PORTION Of THE INTRANARIAL PROCESS (= MEDIAL LAMINA) (SEE HILLENIUS 2000; SIDOR 2003) IS MISSING ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). THE RIGHT INTRANARIAL PROCESS COVERS THE DORSAL SURfACE Of THE PREMAXILLA IN CAPPA/UFSM 0 0 29. IT IS VENTRALLY CONVEX, DORSALLY CONCAVE, AND EXTENDS TOWARDS THE MIDLINE, SEPARATED fROM THE NARIAL fLOOR, AS IN D. elegans . THE fACIAL (=ASCENDING) PROCESS APPEARS TO bE PROPORTIONALLY LESS DEVELOPED THAN IN E. lunensis AND D. elegans . THE DORSAL PORTION Of THE fACIAL PROCESS RUNS POSTERODORSALLY, CONTACTS THE POSTEROLATERAL ASCENDING PROCESS Of THE PREMAXILLA, AND DELIMITATES THE POSTEROLATERAL bORDER Of THE EXTERNAL NARES. THE OPENING Of THE SEPTOMAXILLARY fORAMEN IS UNCERTAIN.
Maxilla. BOTH ELEMENTS ARE ENTIRELY PRESERVED ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). THEY ARE SUbRECTANGULAR IN LATERAL VIEW AND DORSOVENTRALLY DEEP, A CONDITION fOUND IN OTHER ECTENINIIDS. THE MAXILLA OCCUPIES ALMOST ENTIRELY THE LATERAL SURfACE Of THE PREORbITAL REGION Of THE SkULL (OLIVEIRA et al. 2010). AS IN OTHER EUCYNODONTS, SUCH AS Lumkuia fuzzi HOPSON & KITCHING, 2001 AND C. sanjuanensis , THE MAXILLA REACHES ITS GREATEST HEIGHT AT THE LEVEL Of THE CANINE ROOT (MARTÍNEz et al. 2013). ALSO, THE DORSAL MARGIN Of THE MAXILLA IS ANTEROPOSTERIORLY SHORTER IN COMPARISON TO THE VENTRAL MARGIN ( FIG. 4 View FIGURE 4 ), AS IN E. lunensis AND THE HOLOTYPE Of T. riograndensis , WHEREAS IN D. elegans IT IS LESS PRONOUNCED. AS IN OTHER ECTENINIIDS, CAPPA/UFSM 0 0 29 PRESENTS TWO ANTEROPOSTERIORLY ALIGNED INfRAORbITAL fORAMINA IN THE MIDDLE REGION Of THE MAXILLA, HOWEVER, IN E. lunensis AND D. elegans , A THIRD INfRAORbITAL fORAMEN IS OPENED LATERALLY AT THE POINT WHERE LACRIMAL, jUGAL AND MAXILLA CONTACT (MARTÍNEz et al. 1996; 2013). THE POSTERIOR PORTION Of THE MAXILLA CONTACTS THE jUGAL POSTEROVENTRALLY, fORMING THE ANTERIOR ROOT Of THE zYGOMATIC ARCH, THE LACRIMAL POSTERODORSALLY, AND THE NASAL MARGIN DORSALLY ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). THE ANTERODORSAL MARGIN Of THE MAXILLA RUNS ALONG THE NASAL AND MEETS THE EXTERNAL DORSAL OPENING Of THE PARACANINE fOSSA AT THE LEVEL Of THE UPPER CANINE ROOT. COMPARISON Of THE TWO SIDES DEMONSTRATES THAT THE DORSAL OPENING Of THE PARACANINE fOSSAE IS SLIGHTLY DEfORMED. THIS STRUCTURE IS bETTER PRESERVED ON THE LEfT SIDE. THE RIGHT EDGES Of THE PARACANINE fOSSA MEASURE 6 MM ANTEROPOSTERIORLY AND 13 MM DORSOVENTRALLY ( FIG. 2 View FIGURE 2 ). CONVERSELY, THE LEfT PARACANINE fOSSA MEASURES 6 MM ANTEROPOSTERIORLY AND 8 MM DORSOVENTRALLY ( FIG. 3 View FIGURE 3 ). IN CONTRAST TO D. elegans , IN CAPPA/UFSM 0 0 29, UFRGS PV- 1051-T, AND E. lunensis THE PARACANINE fOSSAE ARE OPEN DORSALLY ( FIGS. 4 View FIGURE 4 , 8 View FIGURE 8 ), WHICH ALLOWS THE TIPS Of THE LOWER CANINES TO PROTRUDE THROUGH THE ROOf Of THE SNOUT (MARTÍNEz et al. 2013).
Nasal. CAPPA/UFSM 0 0 29 PRESERVES bOTH NASALS ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). THEY COMPRISE APPROXIMATELY 44% Of THE TOTAL SkULL LENGTH. MEDIALLY, THE NASALS CONTACT EACH OTHER IN A STRAIGHT SUTURE. ON ITS ANTERIOR HALf, THERE IS A SMALL ELEVATION (ANTEROPOSTERIOR CONVEXITY) CLOSE TO THE DORSAL PREMAXILLARY bORDER. IN THE HOLOTYPE Of T. riograndensis AND E. lunensis , THIS REGION IS RELATIVELY fLAT IN COMPARISON WITH ITS REMAINING DORSAL SURfACE. IN D. elegans , THE ENTIRE DORSAL PROfILE Of THE NASAL IS fLAT IN LATERAL VIEW. IT IS PLAUSIbLE THAT THIS DORSAL CONVEXITY IN CAPPA/UFSM 0 0 29 IS AN ARTIfACT Of PRESERVATION bECAUSE THIS REGION IS EXPANDED bY DIAGENESIS ( FIG. 4 View FIGURE 4 ). AS IN OTHER EUCYNODONTS (E.G., HOLOTYPE Of T. riograndensis , Probainognathus jenseni , Chiniquodon theotonicus VON HUENE, 1936), IN DORSAL VIEW THE NASAL IS CURVED ON THE LEVEL Of THE POSTCANINE CONSTRICTION, WHEREAS IN D. elegans IT GENTLY TAPERS fROM THE ANTERIOR END TO THE LEVEL Of THE POSTCANINE CONSTRICTION (MARTÍNEz et al. 2013). LATERALLY, THE NASAL CONTACTS THE MAXILLA ( FIGS. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). ALSO, IT EXPANDS POSTEROLATERALLY CLOSE TO THE LEVEL Of THE ANTERIOR bORDER Of THE ORbIT, SO THAT ITS POSTEROMEDIAL bORDER CONTACTS THE ANTERIOR MARGIN Of THE fRONTAL. ITS POSTEROLATERAL bORDER CONTACTS THE ANTEROMEDIAL MARGIN Of THE PREfRONTAL AND THE ANTEROMEDIAL MARGIN Of THE LACRIMAL. IN D. elegans THE NASAL SLIGHTLY CONTACTS THE PREfRONTAL, IN CONTRAST TO THE CONDITION IN CAPPA/UFSM 0 0 29, UFRGS PV-1051-T, AND E. lunensis , WHERE THE CONTACT bETWEEN THESE bONES IS EXTENSIVE.
Lacrimal. BOTH ELEMENTS ARE ALMOST COMPLETE, LACkING jUST A SMALL fRAGMENT Of THE POSTEROVENTRAL END Of THE RIGHT ELEMENT ( FIGS. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). AS IN THE OTHER ECTENINIIDS, THE LACRIMAL IS ANTEROPOSTERIORLY SHORT, IN CONTRAST TO C. theotonicus , IN WHICH THE LACRIMAL IS ANTERIORLY DEVELOPED, WITH A TRIANGULAR SHAPE IN LATERAL VIEW. THE ANTERIOR LACRIMAL MARGIN IS ROUNDED, fORMING THE ANTERIOR AND VENTRAL ORbITAL RIM. POSTERODORSALLY, THE LACRIMAL CONTACTS THE PREfRONTAL AND THE ANTEROVENTRAL PORTION Of THE jUGAL ( FIGS. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). THE ANTERODORSAL MARGIN Of THE LACRIMAL CONTACTS THE NASAL, WHEREAS ITS ANTEROVENTRAL PORTION CONTACTS THE MAXILLA. IN CAPPA/UFSM 0 0 29, AS IN THE HOLOTYPE Of T. riograndensis , THE LACRIMAL IS REDUCED, bECAUSE IT DOES NOT PRESENT A POSTERIORLY DIRECTED VENTRAL PROCESS THAT fORMS A PRONOUNCED RIDGE IN THE ORbITAL MARGIN, A CONDITION fOUND IN D. elegans , E. lunensis (MARTÍNEz et al. 2013), AND ICTIDOSAURS (E.G., Riograndia guaibensis BONAPARTE et al. 2001; SOARES et al. 2011). THE LACRIMAL Of D. elegans IS DORSOVENTRALLY MORE EXPANDED THAN THAT IN OTHER ECTENINIIDS. THUS, THIS ELEMENT POSSIbLY POSSESSES A LARGE DORSOLATERAL CONTACT WITH THE NASAL AND SIGNIfICANT PARTICIPATION IN THE ORbITAL MARGIN.
Prefrontal. BOTH ELEMENTS ARE ENTIRELY PRESERVED IN CAPPA/UFSM 0029; THEY ARE RECTANGULAR AND POSTEROMEDIALLY EXPANDED, WHERE THEY CONTACT THE fRONTALS ( FIGS. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). IN DORSAL VIEW, THE PREfRONTALS ARE SMALLER THAN THOSE Of TRAVERSODONTIDS ( FIG. 4 View FIGURE 4 ) (E.G., Exaeretodon riograndensis ), IN WHICH THIS bONE IS WIDE AND qUADRANGULAR bONE (AbDALA et al. 2002). CONVERSELY, IN D. elegans AND E. lunensis THE PREfRONTALS ARE NARROWER THAN CAPPA/UFSM 0 0 29 AND UFRGS PV-1051-T AND LACk A POSTEROMEDIAL EXPANSION. THE PREfRONTAL fORMS MOST Of THE DORSAL bORDER Of THE ORbITAL RIM. ANTEROMEDIALLY, THE PREfRONTAL CONTACTS THE POSTEROLATERAL PORTION Of THE NASAL AND ANTEROLATERALLY THE DORSAL MARGIN Of THE LACRIMAL, AS IN THE HOLOTYPE Of T. riograndensis AND E. lunensis , bUT DIffERENTLY fROM D. elegans IN WHICH THE bONE IS MORE ANTERIORLY EXPANDED fORMING A TINY ANTEROMEDIAL CONTACT WITH THE NASAL AND THE LACRIMAL (MARTÍNEz et al. 2013). MEDIALLY, IT CONTACTS THE LATERAL bORDER Of THE fRONTAL AND, POSTERIORLY, THE ANTERODORSAL MARGIN Of THE POSTORbITAL.
Frontal. BOTH ELEMENTS ARE COMPLETE AND PLACED IN THE CENTRAL PORTION Of THE SkULL ROOf ( FIG. 4 View FIGURE 4 ). TOGETHER, THEY HAVE THE SHAPE Of LOzENGE IN DORSAL VIEW, WITH A POSTERIORLY PROjECTED WEDGED PORTION. THE fRONTALS ARTICULATE AGAINST EACH OTHER, fORMING A SMALL MIDLINE CREST THAT TAPERS POSTERIORLY, CONTACTING THE PARIETALS ( FIG. 4 View FIGURE 4 ). IN UFRGS PV-1051-T AND E. lunensis , THE fRONTALS fORM AN AXIAL, fLAT CREST, AND IN D. elegans THE SUTURE bETWEEN fRONTALS IS NOT VISIbLE DUE TO POOR PRESERVATION. THE ANTEROPOSTERIOR LENGTH Of THE fRONTALS IS LARGER THAN THE TRANSVERSE WIDTH. IN CAPPA/UFSM 0 0 29, THE fRONTALS HAVE A LONG POSTEROLATERAL CONTACT WITH THE POSTORbITALS AND A COMPARATIVELY SHORTER ANTEROLATERAL CONTACT WITH THE PREfRONTAL bONES ( FIG. 4 View FIGURE 4 ). THEY ALSO CONTACT THE NASALS ANTERIORLY AND THE PARIETALS POSTEROLATERALLY. IN DORSAL VIEW, THE fRONTAL HAS TWO DEPRESSIONS IN THE ANTEROLATERAL REGION ( FIG. 4 View FIGURE 4 ), WHICH COULD bE AN INDICATION Of A CIRCULAR fORAMEN AS THIS fEATURE IS PRESENT ON THE POSTEROLATERAL PORTION IN E. lunensis (MARTÍNEz et al. 1996). THE fRONTALS Of CAPPA/UFSM 0 0 29 DO NOT CONTACT THE ORbITAL MARGIN, AS IN MOST NON- MAMMALIAfORM CYNODONTS (E.G., ECTENINIIDS, Probainognathus jenseni , Lumkuia fuzzi , AND Exaeretodon riograndensis ). THE SUTURES IN THE ORbITAL WALL CANNOT bE ObSERVED bECAUSE THE INTERNAL ORbITAL REGION IS DAMAGED. HOWEVER, INSIDE THE ORbIT, THE fRONTALS Of THE HOLOTYPE Of T. riograndensis , E. lunensis , AND D. elegans EXTEND VENTRALLY TO fORM PART Of THE INNER ORbITAL WALL, CONTACTING THE LACRIMAL ANTERIORLY, THE PREfRONTAL LATERODORSALLY, AND THE PALATINE VENTRALLY (MARTÍNEz et al. 1996; 2013).
Parietal. THE PARIETALS Of CAPPA/UFSM 0 0 29 ARE ELONGATE AND fUSED TO EACH OTHER ( FIGS. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). THEY COMPRISE APPROXIMATELY 37% Of THE TOTAL SkULL LENGTH IN DORSAL VIEW AND fORM A TALL SAGITTAL CREST THAT ARISES IMMEDIATELY bEHIND Of THE LEVEL Of THE POSTERIOR ORbITAL MARGIN AND EXTENDS POSTERIORLY TO REACH THE ANTERIOR-MOST PORTION Of THE OCCIPITAL (=LAMbDOIDAL) CREST. ANTERIORLY, THE PARIETALS CONTACT THE POSTEROMEDIAL PORTION Of THE fRONTALS AND POSTORbITALS ( FIG. 4 View FIGURE 4 ). POSTERIORLY, THE PARIETAL CONTACTS THE INTERPARIETAL, WHICH CONTRIbUTES TO THE POSTERIOR PORTION Of THE SAGITTAL CREST AND POSTEROVENTRALLY CONTACTS THE SqUAMOSAL, bUT THERE IS NO CLEAR SIGN Of SUTURES bETWEEN THESE bONES ( FIG. 4 View FIGURE 4 ). THUS, IT IS NOT POSSIbLE TO ObSERVE If THE SqUAMOSAL Of CAPPA/UFSM 0 0 29 CONTRIbUTES TO THE fORMATION Of THE SAGITTAL CREST OR PARTICIPATES ONLY IN THE LATERAL PORTION Of THE OCCIPITAL CREST. IN T. riograndensis (UFRGS PV-1051-T) THE SqUAMOSAL CONTRIbUTES ONLY TO THE LATERAL PORTION Of THE OCCIPITAL CREST. IN CONTRAST, IN E. lunensis AND Probainognathus , THE SqUAMOSAL CONTRIbUTES TO THE POSTEROLATERAL PORTION Of THE SAGITTAL CREST AND POSTERIOR PORTION Of THE OCCIPITAL CREST, MORE EXTENSIVELY DEVELOPED IN THE LATTER TAXON (MARTÍNEz et al. 1996). THE PINEAL fORAMEN IS AbSENT, AS IN OTHER PRObAINOGNATHIANS (HOPSON & KITCHING 2001; LIU & OLSEN 2010).
Postorbital. CAPPA/UFSM 0 0 29 PRESERVES bOTH POSTORbITALS ( FIGS. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 ). DORSALLY THEY PARTICIPATE IN A SMALL PORTION Of THE SkULL ROOf, AND THEIR SUTURAL CONTACTS ARE CLEAR. THE POSTORbITAL bEARS A POSTERIOR PROCESS WITH A SEMICIRCULAR SHAPE, WHICH EXTENDS POSTERIORLY TOWARDS THE SkULL MIDLINE. THE POSTERIOR PROCESS bROADLY OVERLAPS THE POSTEROLATERAL PORTION Of THE fRONTAL AND THE ANTEROMEDIAL MARGIN Of THE PARIETAL. ANTERIORLY, THE POSTORbITAL ALSO CONTACTS THE POSTERIOR bORDER Of THE PREfRONTAL. THE POSTORbITAL bAR fORMS bOTH POSTERODORSAL AND POSTERIOR ORbITAL MARGIN. IT IS THIN AND SUbCIRCULAR IN CROSS-SECTION, AS IN THE HOLOTYPE Of T. riograndensis , E. lunensis , AND Chiniquodon sanjuanensis (MARTÍNEz & FORSTER 1996; MARTÍNEz et al. 1996; OLIVEIRA et al. 2010). IN D. elegans , THIS CONDITION IS UNkNOWN bECAUSE bOTH THE LEfT POSTORbITAL AND POSTERIOR PORTION Of THE RIGHT ONE ARE NOT PRESERVED. THE POSTORbITAL bAR Of CAPPA/UFSM 0 0 29 CONTRIbUTES A DESCENDING PROCESS, WHICH ANTERIORLY OVERLAPS THE ASCENDING PROCESS Of THE jUGAL ( FIG. 3 View FIGURE 3 ), A CONDITION SHARED WITH THE HOLOTYPE Of T. riograndensis AND E. lunensis (PVSJ 481). ON THE OTHER HAND, THE HOLOTYPE Of E. lunensis (PVSJ 422) DOES NOT PRESERVE THIS REGION. ACCORDING TO OLIVEIRA et al. (2010), T. riograndensis (UFRGS PV-1051-T) PRESENTS A POSSIbLE AUTAPOMORPHIC fEATURE IN THE POSTORbITAL: IT OVERLAPS THE jUGAL IN THE VENTRAL MARGIN Of THE ORbIT. HOWEVER, THIS fEATURE IS NOT AN AUTAPOMORPHY Of HOLOTYPE UFRGS PV-1051-T bUT, INSTEAD, A CONDITION SHARED WITH E. lunensis (PVSJ 481). IN D. elegans THIS fEATURE IS UNkNOWN.
Jugal. BOTH SIDES Of THE SPECIMEN PARTIALLY PRESERVE THIS bONE ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). A SMALL ANTERIOR PORTION THAT CONTACTS THE LACRIMAL ANTERODORSALLY AND THE MAXILLA ANTERIORLY IS DISPLACED fROM THE RIGHT ELEMENT. IN ADDITION, bOTH POSTERIOR PORTIONS Of THE zYGOMATIC ARCH ARE fRACTURED, AND THE RIGHT ARCH IS bETTER PRESERVED THAN THE LEfT ONE, WHICH IS bROkEN ON ITS POSTERIOR END. THE jUGAL LARGELY CONTRIbUTES TO THE zYGOMATIC ARCH, bUT DUE TO bAD PRESERVATION, THE POSTERIOR CONTACT Of THE jUGAL AND SqUAMOSAL IS UNCERTAIN. COMPARED TO OTHER NON-MAMMALIAfORM CYNODONTS, THE zYGOMATIC ARCH Of CAPPA/UFSM 0 0 29 IS NARROW AND SLENDER, AS IN UFRGS PV-1051-T AND THE PRESERVED ANTERIOR PORTIONS Of THE jUGAL Of D. elegans AND E. lunensis (MARTÍNEz et al. 1996; 2013; OLIVEIRA et al. 2010). ON THE OTHER HAND, THE zYGOMATIC ARCH Of CHINIqUODONTIDS, Probainognathus jenseni , AND OTHER CYNOGNATHIANS (E.G., Exaeretodon riograndensis ) IS MORE RObUST. THE jUGAL fORMS APPROXIMATELY 50% Of THE POSTEROVENTRAL ORbITAL RIM ( FIG. 2 View FIGURE 2 ). DORSALLY THE LATERAL PORTION Of THE POSTORbITAL, IN CAPPA/UFSM 0 0 29 AND IN T. riograndensis (UFRGS PV-1051-T) PRESENT A SHORT jUGAL/LACRIMAL CONTACT, IN CONTRAST TO THE CONDITION IN D. elegans AND E. lunensis (SEE MARTÍNEz et al. 1996; 2013). IN THE POSTERIOR PORTION Of THE ORbITAL RIM, THE ASCENDING PROCESS Of THE jUGAL CONTACTS THE DESCENDING PROCESS Of THE POSTORbITAL, WHICH OVERLAPS THE fORMER IN THE ORbITAL MARGIN ( FIG. 3 View FIGURE 3 ), AS IN THE HOLOTYPE Of T. riograndensis (OLIVEIRA et al. 2010) AND E. lunensis (PVSJ 481).
Interparietal. THIS bONE IS LIkELY ObSERVAbLE IN DORSAL, LATERAL, AND OCCIPITAL VIEWS, WITH ONLY A SMALL PORTION Of ITS RIGHT DORSAL EDGE MISSING ( FIGS. 4 View FIGURE 4 , 6 View FIGURE 6 ). HOWEVER, IT IS NOT POSSIbLE TO ObSERVE ITS SUTURES WITH THE OTHER bONES, AND SO ITS LIMITS MUST REMAIN CONjECTURAL. IN DORSAL VIEW, THE INTERPARIETAL Of CAPPA/UFSM 0 0 29 CONTRIbUTES TO THE POSTERIOR END Of THE SAGITTAL CREST AND PARTICIPATES IN THE POSTERODORSAL PORTION Of THE OCCIPITAL CREST, AS THE HOLOTYPE UFRGS PV-1051-T (SEE OLIVEIRA et al. 2010). AS IN T. riograndensis (UFRGS PV-1051-T) AND E. lunensis , TWO CONCAVITIES ARE VISIbLE IN OCCIPITAL VIEW, IN THE AREA SUPPOSEDLY CLOSE TO THE CONTACT WITH THE TAbULARS.
Squamosal. THE SqUAMOSAL Of CAPPA /UFSM 0 0 29 IS POORLY PRESERVED, AND THE SUTURES THAT DELIMIT THIS bONE ARE NOT ObSERVAbLE ( FIGS. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 6–7 View FIGURE 6 View FIGURE 7 ). THIS ELEMENT IS SIGNIfICANTLY DAMAGED ON bOTH SIDES, AND THE zYGOMATIC ARCHES ARE fRAGMENTED ON THE CONTACT bETWEEN SqUAMOSAL AND jUGAL ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). THE LEfT SqUAMOSAL IS bETTER PRESERVED THAN THE RIGHT ONE ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). THE AREA Of ARTICULATION Of THE SkULL WITH THE jAW IS DAMAGED. HOWEVER, THERE IS A SMALL ELEMENT THAT MIGHT CORRESPOND TO qUADRATE/qUADRATOjUGAL ( FIG. 3 View FIGURE 3 ).
THE SqUAMOSAL ALSO fORMS THE ANTEROVENTRAL PORTION Of THE OCCIPITAL CREST AND A SMALL VENTRAL PART Of THE POSTERIOR REGION Of THE SkULL ( FIGS. 4 View FIGURE 4 , 7 View FIGURE 7 ). IN CAPPA / UFSM 0 0 29 AND IN THE HOLOTYPE Of T. riograndensis , THE PTERYGOPAROCCIPITAL fORAMEN OPENS ANTERIORLY INTO THE TEMPORAL fOSSA. IN E. lunensis THIS fORAMEN IS CLOSED bY THE qUADRATE RAMUS Of THE EPIPTERYGOID . DUE TO THE STATE Of PRESERVATION Of CAPPA / UFSM 0 0 29 IN THIS REGION, THE VISUALIzATION Of THE SUTURES IS NOT COMPLETELY POSSIbLE . IN THE LATERAL SURfACE Of THE SqUAMOSAL , THERE IS A DORSOVENTRAL SULCUS (=EXTERNAL AUDITORY MEATUS) ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ) AND, AT THE POSTERIOR END, THIS bONE PARTICIPATES IN THE OCCIPITAL CREST. THE SqUAMOSAL ALSO fORMS THE POSTERIOR PORTION Of THE LATERAL WALL Of THE OCCIPITAL CREST, IN OCCIPITAL VIEW ( FIG. 6 View FIGURE 6 ) . THE SqUAMOSAL EXTENSIVELY CONTACTS THE TAbULAR AND THESE bONES DELIMITATE THE POSTERIOR OPENING Of THE POST-TEMPORAL fORAMEN, AS IN T. riograndensis ( UFRGS PV-1051 -T) (OLIVEIRA et al. 2010) . CONVERSELY, IN E. lunensis THIS OPENING IS SURROUNDED ENTIRELY bY THE TAbULAR bONE.
UNfORTUNATELY, THE VISUALIzATION Of THE SUTURES Of THE SqUAMOSAL WITH THE SURROUNDING bONES IS IMPOSSIbLE IN CAPPA/UFSM 0 0 29, PREVENTING THE ObSERVATION If IT CONTRIbUTES ONLY TO THE POSTERIOR PORTION Of THE LATERAL WALL Of THE OCCIPITAL CREST, AS IN THE HOLOTYPE UFRGS PV-1051-T, OR If IT ALSO CONTRIbUTES TO THE POSTEROLATERAL PORTION Of THE SAGITTAL CREST, AS IN E. lunensis AND Probainognathus (MARTÍNEz et al. 1996).
Tabular. CAPPA/UFSM 0 0 29 PRESERVES bOTH TAbULARS ( FIG. 6 View FIGURE 6 ). THEY fORM THE DORSOLATERAL PART Of THE OCCIPUT AND, TOGETHER WITH THE SqUAMOSAL, CONTRIbUTE TO THE DORSOLATERAL AND LATERAL MARGINS Of THE OCCIPITAL CREST, AS IN T. riograndensis (UFRGS PV-1051-T) AND E. lunensis (MARTÍNEz et al. 1996; OLIVEIRA et al. 2010). THE TAbULAR CONTACTS THE INTERPARIETAL DORSOMEDIALLY AND THE SqUAMOSAL LATERALLY, bUT THE EXACT SUTURAL CONTACTS WITH THE SURROUNDING ELEMENTS ARE NOT EVIDENT.
Supraoccipital. IN OCCIPITAL VIEW, THE SUPRAOCCIPITAL Of CAPPA / UFSM 0 0 29 DELIMITS ALMOST THE ENTIRE DORSAL MARGIN Of THE fORAMEN MAGNUM . THIS ELEMENT CONTACTS THE INTERPARIETAL DORSALLY, AND THE TAbULARS LATERALLY ( FIG. 6 View FIGURE 6 ). THE CONTACT WITH THE EXOCCIPITAL IS ObSCURED bY INCRUSTATION. THE fORAMEN MAGNUM IS RELATIVELY LARGE AND TRIANGULAR IN OUTLINE ( FIG. 6 View FIGURE 6 ). IN E. lunensis , THE fORAMEN MAGNUM IS OVAL-SHAPED, WHEREAS IN UFRGS PV-1051-T IT IS MODIfIED bY THE DIAGENESIS. THE VENTRAL MARGIN Of THE fORAMEN MAGNUM Of CAPPA/UFSM 0 0 29 ( FIG. 7 View FIGURE 7 ), IN bOTH THE POSTERIOR PORTION Of THE PARAbASISPHENOID COMPLEX AND ALMOST ALL bASIOCCIPITAL, ARE EITHER MISSING OR SUffERED DAMAGES, EXPOSING AN INTERNAL PORTION Of THE bRAINCASE.
Exoccipital. BOTH EXOCCIPITAL bONES ARE COMPLETE, LOCATED VENTROLATERALLY TO THE fORAMEN MAGNUM. THE EXOCCIPITALS ARE ELONGATED, AND THEIR EXTREMITIES fORM bULbOUS OCCIPITAL CONDYLES. AS WELL AS IN OTHER ECTENINIIDS, THE EXOCCIPITAL fORM ENTIRELY THE OCCIPITAL CONDYLES. THESE ELEMENTS CANNOT bE ObSERVED IN DORSAL VIEW, AS THEY ARE COVERED bY THE POSTERIORLY EXPANDED OCCIPITAL CRESTS, SUCH AS IN UFRGS PV-1051-T AND E. lunensis . THIS fEATURE DIffERS fROM THE ObSERVED CONDITION IN NON-ECTENINIID EUCYNODONTS, IN WHICH THEY ARE EXPOSED IN DORSAL VIEW [E.G., Lumkuia fuzzi , Massetognathus ochagaviae BARbERENA, 1981 (UFRGS PV-0255-T)]. THE OCCIPITAL CONDYLES DELIMIT THE fORAMEN MAGNUM IN THEIR LATERAL AND VENTRAL MARGINS. THE EXOCCIPITAL CONTACTS THE SUPRAOCCIPITAL DORSALLY, THE PAROCCIPITAL PROCESS Of THE OPISTHOTIC LATERALLY, AND THE PRESERVED PORTION Of THE bASIOCCIPITAL MEDIOVENTRALLY. BELOW THE CONTACT WITH THE PRESERVED PORTION Of THE bASIOCCIPITAL, THIS LATER ELEMENT ANTERIORLY CONTACTS THE PARAbASISPHENOID COMPLEX, WHICH IS VISIbLE VENTRALLY. IN CAPPA/UFSM 0 0 29 AND UFRGS PV-1051-T, THE HYPOGLOSSAL fORAMEN IS NOT VISIbLE, bUT THE jUGULAR fORAMEN IS PRESENT IN THE SECOND SPECIMEN. IN E. lunensis , A SMALL HYPOGLOSSAL fORAMEN IS LOCATED bELOW THE OCCIPITAL CONDYLES AND POSTEROMEDIALLY TO THE jUGULAR fORAMEN (MARTÍNEz et al. 1996).
Pterygoid. DESPITE THE DEfICIENT PRESERVATION Of THE VENTRAL ASPECT Of THE NEW SPECIMEN, THE PTERYGOID IS ALMOST COMPLETE bUT SIGNIfICANTLY AffECTED bY TAPHONOMIC DEfORMATION ( FIG. 7 View FIGURE 7 ). ANTERIOR TO THE PTERYGOID, THE bONES Of THE SECONDARY PALATE ARE MISSING. THE POSTERIOR PORTION Of THE PTERYGOID IS bROkEN ON THE CONTACT WITH THE PARAbASISPHENOID COMPLEX, WHERE VISUALIzATION Of SUTURES IS NOT POSSIbLE GIVEN THE POOR PRESERVATION Of THIS REGION ( FIG. 7 View FIGURE 7 ). THE PTERYGOID fLANGES ARE LONG, NARROW, SUbTRIANGULAR IN VENTRAL VIEW, AND TAPER VENTRALLY. ON THE OTHER HAND, IN UFRGS PV-1051-T THEY ARE MORE EXPANDED LATERALLY AND STOUT, WITH A TRIANGULAR-SHAPED CROSS SECTION (OLIVEIRA et al. 2010). IN E. lunensis , THE PTERYGOID fLANGES ARE qUADRANGULAR, bROAD AND RObUST, EXTENDED bACk bEHIND THE ORbITS, AS THEIR TIPS CURVE POSTEROMEDIALLY (MARTÍNEz et al. 1996). THE PTERYGOID Of E. lunensis fORMS THREE RIDGES, ONE MEDIAN AND TWO LATERAL, RADIATING fORWARD fROM A POINT AT THE fRONT Of THE bASISPHENOID (MARTÍNEz et al. 1996). IN THE NEW SPECIMEN AND THE HOLOTYPE Of T. riograndensis , THIS REGION IS DAMAGED, WHICH PREVENTS CONfIRMATION Of SUCH RIDGES.
Parabasisphenoid complex. TAPHONOMIC DEfORMATION STRONGLY AffECTED THE PARAbASISPHENOID COMPLEX Of CAPPA/UFSM 0 0 29 ( FIG. 7 View FIGURE 7 ). THE COMPLEX IS PLACED IN THE MEDIAL PORTION Of THE bASICRANIUM AND HAS AN ELONGATED TRIANGULAR SHAPE. IN THE ANTERIOR PORTION, THE bASISPHENOID IS bROkEN CLOSE TO THE CONTACT WITH THE PTERYGOIDS AND ITS ALMOST ENTIRE POSTERIOR PORTION IS MISSING OR DAMAGED. IN THIS REGION, DUE TO ITS POOR PRESERVATION, IT IS NOT POSSIbLE TO ObSERVE THE CONTACT Of THE bASIOCCIPITAL WITH THE POSTERIOR PORTION Of THE bASICRANIUM AND TO THE VENTRAL RIM Of THE fORAMEN MAGNUM. IN CAPPA/UFSM 0 0 29 AND T. riograndensis (UFRGS PV-1051-T) THE PARAbASISPHENOID COMPLEX PRESENTS A LOW MEDIAN PARAbASISPHENOID RIDGE AND TWO WELL-DEVELOPED bASISPHENOID WINGS LATERALLY (OLIVEIRA et al. 2010). IN THE MEDIAL PORTION Of THIS ELEMENT, THERE IS A fRACTURE THAT ObSCURES DETAILS Of THIS REGION IN CAPPA/UFSM 0 0 29. IN THE NEW SPECIMEN, IN THE HOLOTYPE Of T. riograndensis , AND IN E. lunensis , THE PARAbASISPHENOID LACkS fORAMINA fOR THE INTERNAL CAROTIDS (MARTÍNEz et al. 1996; OLIVEIRA et al. 2010).
Prootic and Epipterygoid. BOTH ELEMENTS ARE TAPHONOMICALLY AffECTED, AND THEIR SUTURES ARE UNCERTAIN. THE PROOTIC IS ObSERVAbLE IN LATERAL AND VENTRAL VIEWS ( FIGS. 6–7 View FIGURE 6 View FIGURE 7 ), ALTHOUGH THE CONTACTS WITH OTHER bONES ARE NOT VISIbLE. A LARGE AND ROUND POST-TEMPORAL fORAMEN IS ANTEROVENTRALLY OPENED, bETWEEN THE PROOTIC AND EPIPTERYGOID ( FIG. 6 View FIGURE 6 ). IN VENTRAL VIEW, THE PROOTIC CONTACTS THE PARAbASISPHENOID COMPLEX MEDIALLY AND THE SqUAMOSAL POSTERIORLY. THE EPIPTERYGOID Of CAPPA / UFSM 0 0 29 IS VISIbLE IN LATERAL VIEW .
Mandible. THE LOWER jAW PRESERVES THE ANTERIOR AND MIDDLE PORTIONS Of bOTH DENTARIES, WHEREAS POSTERODORSALLY IT IS DAMAGED IN SOME DEGREE ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). THE MANDIbULAR SYMPHYSIS IS fUSED, ANTEROPOSTERIORLY SLOPED AND DORSOVENTRALLY CONVEX ON ITS ANTERIOR PORTION. THE ANGULAR AND CORONOID PROCESSES Of CAPPA /UFSM 0 0 29 ARE POSTERIORLY DAMAGED. ITS POSTDENTARY ELEMENTS ARE PARTIALLY PRESERVED. THE LOWER jAW Of T. riograndensis ( HOLOTYPE UFRGS PV-1051-T) IS bETTER PRESERVED THAN IN CAPPA /UFSM 0 0 29, SO THE fORMER SPECIMEN PROVIDES MORE INfORMATION THE AbOUT POSTDENTARY bONES AND LOWER DENTITION.
THE DENTARY IS LONG, DEEP, AND RObUST ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). AS ALREADY MENTIONED, THE DENTARIES ARE fUSED AT THE MANDIbULAR SYMPHYSIS, WHICH POSTERIORLY EXTENDS TO THE ANTERIOR LEVEL Of THE UPPER CANINE TOOTH . THE fUSED SYMPHYSIS IS TYPICAL fOR MOST NON-MAMMALIAMORPH EUCYNODONTS, EXCEPT IN MORE DERIVED fORMS (SEE BONAPARTE 1980; BONAPARTE et al. 2001; MARTINELLI et al. 2016b). THE bODY Of THE DENTARY IS ALMOST UNIfORMLY DEEP ALONG ITS LENGTH ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ), AS IN THE HOLOTYPE Of T. riograndensis (SEE OLIVEIRA et al. 2010).
THE POSTERODORSAL PORTIONS Of bOTH CORONOID PROCESSES ARE DAMAGED, bUT THE LEfT DENTARY IS bETTER PRESERVED THAN THE RIGHT ONE ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). THE CORONOID PROCESS Of CAPPA/UFSM 0 0 29 IS bROAD AND DEEP, POSTERIORLY EXTENDING bEYOND THE LEVEL Of THE ANGULAR PROCESS, AS IN THE HOLOTYPE Of T. riograndensis (SEE OLIVEIRA et al. 2010). IN COMPARISON, THE CORONOID PROCESS Of E. lunensis IS THIN AND TALL, WITH ITS ANTERIOR MARGIN SLIGHTLY CONCAVE, AND ITS DORSAL bORDER IS REACHING THE TOP Of THE SAGITTAL CREST WHEN ARTICULATED TO THE SkULL (MARTÍNEz et al. 1996). IN Chiniquodon sanjuanensis (PVSJ 411), THIS STRUCTURE IS ALSO TALL AND REACHES THE LEVEL Of THE SAGITTAL CREST (MARTÍNEz & FORSTER 1996). IN MOST GOMPHODONTS, SUCH AS THE TRAVERSODONTID Exaeretodon riograndensis , THE CORONOID PROCESS IS CONSIDERAbLY DEVELOPED AND TALLER THAN IN PRObAINOGNATHIANS (LIPARINI et al. 2013).
A SHALLOW AND bROAD MASSETERIC fOSSA EXTENDS ANTEROPOSTERIORLY ON THE LATERAL RAMUS Of THE DENTARY ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). IN CAPPA/UFSM 0 0 29, UFRGS PV-1051-T, AND E. lunensis (PVSJ 481), THE MASSETERIC fOSSA OCCUPIES ALMOST ENTIRELY THE LATERAL SURfACE Of THE POSTERIOR HALf Of DENTARY (SEE OLIVEIRA et al. 2010). IN COMPARISON, IN THE HOLOTYPE Of E. lunensis (PVSJ 422), IT IS SLIGHTLY LESS WIDE (MARTÍNEz et al. 1996). IN THE NEW SPECIMEN, THIS STRUCTURE bEGINS bEHIND THE LAST POSTCANINE, AS IN THE HOLOTYPE Of T. riograndensis (OLIVEIRA et al. 2010). IN OTHER CYNODONT GROUPS, SUCH AS IN Diademodon tetragonus SEELEY, 1894 , THE MASSETERIC fOSSA EXTENDS WELL fORWARD, UNTIL THE LEVEL Of THE SIXTH LOWER POSTCANINE (MARTINELLI et al. 2009) AND IN Chaliminia musteloides BONAPARTE, 1980 THE MASSETERIC fOSSA IS bROAD, DEVELOPED ON THE ENTIRE LATERAL SURfACE Of THE CORONOID PROCESS, AND ANTERIORLY EXTENDED TO THE LEVEL Of THE LAST UPPER POSTCANINE (MARTINELLI & ROUGIER 2007).
THE LATERAL SURfACE Of THE ANTERIOR REGION Of THE DENTARY Of CAPPA/UFSM 0 0 29 IS PERfORATED bY THE MENTAL fORAMEN, WHICH IS LOCATED AT THE LEVEL Of THE fIRST UPPER POSTCANINES ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). IN THE HOLOTYPE Of T. riograndensis , THIS fORAMEN IS CLOSE TO THE LEVEL Of THE THIRD POSTCANINE (SEE OLIVEIRA et al. 2010). E. lunensis PRESENTS TWO fORAMINA, THE fIRST ONE LARGER, LOCATED IMMEDIATELY bEHIND THE UPPER CANINE AND A SECOND ONE SMALLER AND DORSOLATERALLY PLACED bENEATH THE THIRD POSTCANINE (MARTÍNEz et al. 1996). THE MECkELIAN GROOVE Of CAPPA/UFSM 0 0 29 IS NOT VISIbLE.
THE TIP Of THE ANGULAR PROCESS LACkS IN bOTH DENTARIES Of CAPPA / UFSM 0 0 29. IN THE HOLOTYPE UFRGS PV- 1051 - T, THE ANGULAR PROCESS HAS A POSTERIORLY PROjECTED TIP (OLIVEIRA et al. 2010). IN MOST Of THE TRAVERSODONTIDS THIS OfTEN PROCESS IS PROMINENT AND ACUTE (LIPARINI et al. 2013). IN CAPPA / UFSM 0 0 29 , THE DENTARY ENDS NEAR THE jAW ARTICULATION, AS IN T. riograndensis ( UFRGS PV-1051 -T).
ON bOTH SIDES Of THE LOWER jAW, THE POSTDENTARY bONES ARE PARTIALLY PRESERVED, COMPRISING A fRAGILE STRUCTURE IN THE POSTERIOR END Of THE DENTARY ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 , 7 View FIGURE 7 ). THEY ARE LOCATED MEDIALLY AND POSTERODORSALLY TO THE ANGULAR PROCESS, AND VENTRALLY TO THE LEVEL Of THE PRESERVED PORTIONS Of THE CORONOID PROCESS. THE POSTDENTARY bONES ARE NOT READILY IDENTIfIAbLE bECAUSE Of THE DEfICIENT PRESERVATION, AND THEIR CONTACTS ARE NOT CLEAR. THE LEfT ANGULAR, SURANGULAR, AND PREARTICULAR ARE PRESERVED, WHEREAS ONLY PARTS Of THE RIGHT ANGULAR AND SURANGULAR ARE IDENTIfIAbLE. IN VENTRAL VIEW, bOTH RIGHT AND LEfT ANGULAR bONES ARE PARTIALLY VISIbLE. THE ANGULAR ARTICULATES bETWEEN THE SURANGULAR AND PREARTICULAR, DORSALLY CONTACTING THE PRESERVED PORTION Of THE SURANGULAR AND, LATEROVENTRALLY, THE PRESERVED PORTION fROM PREARTICULAR. THE ANGULAR fORMS AN ANTEROPOSTERIORLY LONG bAR, WHICH EXTENDS MEDIALLY AND POSTERODORSALLY TO THE LEVEL Of THE ANGULAR PROCESS. THE ANGULAR AND SURANGULAR ARE APPARENTLY UNfUSED, AND A PUTATIVE CONTACT IS ObSERVAbLE bETWEEN THESE bONES, AS IN THE HOLOTYPE Of T. riograndensis ( UFRGS PV-1051 -T) (SEE OLIVEIRA et al. 2010) ( FIGS. 3 View FIGURE 3 , 7 View FIGURE 7 ). IN UFRGS PV-1051 - T, THE POSTDENTARY bONES fORM A LONG AND SLENDER bAR THAT EXTENDS bELOW THE MEDIAL CREST Of THE DENTARY (OLIVEIRA et al. 2010). THE ANGULAR PLUS SURANGULAR Of THE HOLOTYPE Of T. riograndensis ARE PARTIALLY PARALLEL TO EACH OTHER, EXTENDED MEDIALLY bEHIND THE ANGULAR PROCESS, AND CONTACT THE PREARTICULAR AND ARTICULAR POSTERODORSALLY.
THE SURANGULAR, VISIbLE ON THE LATERAL VIEW Of bOTH SIDES Of THE LOWER jAW, IS bETTER PRESERVED IN THE LEfT SIDE Of THE SPECIMEN ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). THE ANTERIOR PORTION Of THE SURANGULAR fORMS A bAR WITH TAPERED ENDS WITH AN ANGLE Of AbOUT 45° IN RELATION TO THE CENTRAL AXIS Of THE bODY Of THE DENTARY. THE LEfT PREARTICULAR IS INCOMPLETE, WHEREAS THE RIGHT ONE IS MISSING. IN VENTRAL VIEW, THE CONTACT bETWEEN THE PREARTICULAR AND ANGULAR IS ObSERVAbLE, AS IN T. riograndensis (UFRGS PV-1051-T). UNfORTUNATELY, bOTH ARTICULAR AND RETROARTICULAR PROCESSES ARE NOT PRESERVED.
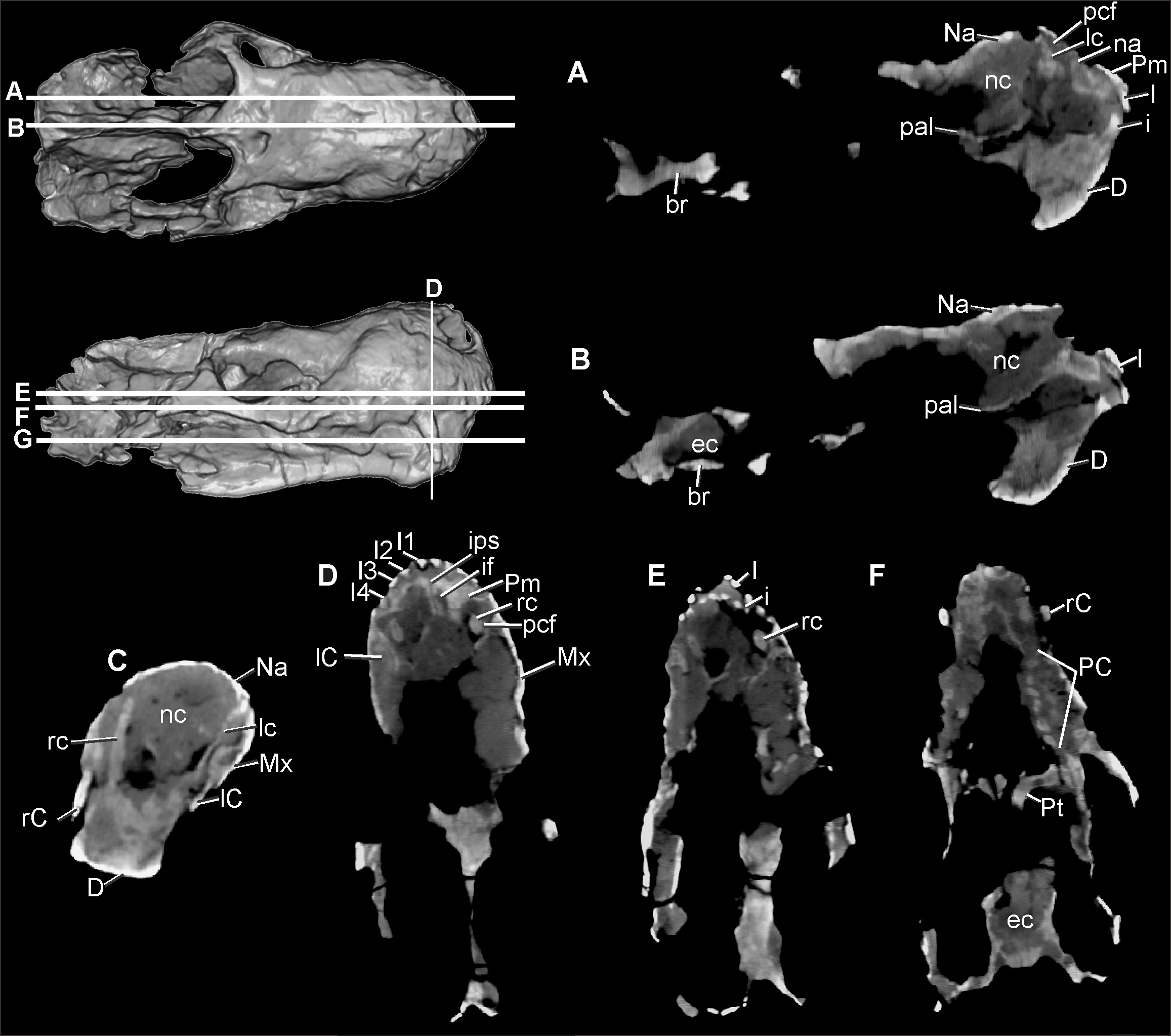
Dentition. AS THE LOWER jAW IS IN OCCLUSION, THE LOWER DENTITION IS NOT EXPOSED ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 , 5 View FIGURE 5 , 8 View FIGURE 8 ). IN CONTRAST TO THE CONDITION IN OTHER ECTENINIIDS, SERRATIONS CANNOT bE ObSERVED IN ANY TOOTH, ALTHOUGH PRObAbLY THIS MAY bE DUE TO POOR PRESERVATION, AS SERRATIONS ARE ObSERVAbLE IN THE HOLOTYPE ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ). THE UPPER LEfT DENTITION PRESENTS fOUR INCISORS ( FIGS. 5 View FIGURE 5 , 8 View FIGURE 8 ), ONE CANINE, AND fIVE POSTCANINES, bUT WITH SPACE fOR UP TO EIGHT POSTCANINE TEETH ( FIG. 3 View FIGURE 3 ). THE UPPER RIGHT DENTITION SHOWS THE SAME NUMbER Of INCISORS AND CANINES, bUT POSTCANINES ARE NOT PRESERVED ( FIG. 2 View FIGURE 2 ). THE UPPER DENTITION Of HOLOTYPE Of T. riograndensis AND E. lunensis PRESENTS 4I /1C/8PC (MARTÍNEz et al. 1996; OLIVEIRA et al. 2010), WHEREAS IT IS 4I /1C/7PC IN D. elegans (MARTÍNEz et al. 2013). HOWEVER, CHANGES IN THE NUMbER Of POSTCANINES ALONG ONTOGENY ARE NOT kNOWN IN ECTENINIIDS DUE TO THE LIMITED SAMPLE Of SPECIMENS, bUT THE DIffERENCE IN POSTCANINE NUMbERS ALONG DIffERENT STAGES IS fREqUENT IN CYNODONTS (E.G., FOURIE 1963).
THE UPPER INCISORS ARE THIN AND LONG ( FIG. 5 View FIGURE 5 ), WITH THE fIRST RIGHT ONE bEING SMALLER THAN THE fOLLOWING AND SUCCESSIVELY LARGER TEETH. THE LEfT SIDE PRESENTS THE SAME CONDITION. THE POSTERIOR INCREASE IN THE SIzE Of THE INCISORS IS SEEN IN THE HOLOTYPE UFRGS PV-1051- T, bUT IT IS DIffERENT fROM E. lunensis ( PVSJ 422 View Materials ), IN WHICH THE fIRST TWO INCISORS ARE THE LONGEST AND THINNEST. THE fOUR INCISORS Of CAPPA / UFSM 0 0 29 ARE IMPLANTED IN THE PREMAXILLA, AND bECAUSE THIS bONE ENTERS MEDIALLY TO THE ANTEROVENTRAL PORTION Of THE MAXILLA, THE fOURTH INCISOR SEEMS TO bE IMPLANTED IN THE LATTER bONE. THE ROOT Of THE fOURTH RIGHT INCISOR Of CAPPA / UFSM 0 0 29 IS EXPOSED DUE TO DAMAGE TO THE MAXILLA. THIS TOOTH, AS WELL AS THE THIRD AND fOURTH LEfT INCISORS, ARE SLIGHTLY DAMAGED IN THEIR TIP.
IN CAPPA / UFSM 0 0 29 AND UFRGS PV-1051 -T (SEE OLIVEIRA et al. 2010), THE THIRD AND fOURTH INCISOR ARE SLIGHTLY MORE SPACED fROM EACH OTHER THAN THE REMAINING ONES, IN CONTRAST TO E. lunensis , IN WHICH THE INCISORS ARE EVENLY SPACED. IN D. elegans THE INCISORS ARE UNEVENLY LOCATED, AND THE TWO POSTERIOR TEETH ARE MORE SPACED IN COMPARISON TO THE ANTERIOR ONES (MARTÍNEz et al. 2013).
CAPPA/UFSM 0 0 29 LACkS VISIbLE SERRATIONS ON THE INCISORS ( FIG. 5 View FIGURE 5 ), ALTHOUGH THIS IS POSSIbLY AN ARTIfACT Of PRESERVATION. IN E. lunensis THE SERRATIONS ARE AbSENT (MARTÍNEz et al. 1996), WHEREAS IN UFRGS PV-1051-T THE DISTAL MARGIN Of THE LAST THREE INCISORS ARE SERRATED AND ON THE LINGUAL SIDE THERE IS A HIGH VERTICAL RIDGE IN THE fOURTH INCISOR (OLIVEIRA et al. 2010). IN THE NEW SPECIMEN, THE LINGUAL SIDE IS NOT VISIbLE. AS IN THE OTHER ECTENINIIDS, THE LAbIAL SURfACE Of THE INCISORS IS fLAT AND POSSESS LONGITUDINAL STRIATIONS, MAINLY IN THE TWO POSTERIOR TEETH, A CONDITION fOUND IN E. lunensis AND D. elegans (MARTÍNEz et al. 1996; 2013).
A SLIGHT PROCUMbENT ORIENTATION Of THE INCISORS IS PRESENT IN CAPPA /UFSM 0 0 29, bUT THIS fEATURE POSSIbLY MAY bE CAUSED bY THE LATERODORSAL COMPRESSION THAT ANTERIORLY DISPLACED THEM fORWARDS. PROCUMbENT INCISORS ARE ALSO PRESENT IN E. lunensis (MARTÍNEz et al. 1996), WHEREAS IN OTHER PRObAINOGNATHIAN CYNODONTS (E.G., UFRGS PV-1051- T, D. elegans , Chiniquodon sanjuanensis , Probainognathus jenseni , Lumkuia fuzzi ) A VERTICAL OR SLIGHTLY RECUMbENT ORIENTATION IS PREVALENT.
IN CAPPA / UFSM 0 0 29 , THE UPPER CANINES ARE LONG, WELL DEVELOPED, LAbIOLINGUALLY COMPRESSED, AND SLIGHTLY RECURVED bACkWARD ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 , 5 View FIGURE 5 , 8 View FIGURE 8 ). BOTH ARE PRACTICALLY COMPLETE, WITH ONLY A SMALL PORTION Of EACH TIP MISSING. AS IN OTHER ECTENINIIDS, THERE IS A DIASTEMA bETWEEN THE LAST INCISOR AND THE CANINE . THE CANINES ARE qUITE LONG AND ALMOST REACH THE VENTRAL MARGIN Of THE DENTARY. THE CROSS-SECTION Of THE CANINE IS ROUND NEAR ITS IMPLANTATION IN THE MAXILLA AND bECOMES MORE OVAL TOWARDS ITS TIP, SUCH AS IN OTHER ECTENINIIDS (MARTÍNEz et al. 1996; 2013).
THE UPPER CANINES LACk VISIbLE SERRATIONS, bUT AS fOR THE INCISORS, THIS IS POSSIbLY AN ARTIfACT Of PRESERVATION . D. elegans LACkS SERRATIONS ON THE PRESERVED fRAGMENTS Of ITS fUNCTIONAL CANINES AS WELL AS ON THE REPLACEMENT CANINES, bUT THESE ARE bADLY PRESERVED AS WELL (MARTÍNEz et al. 2013) . CONVERSELY, IN E. lunensis THE CANINES ARE fINELY SERRATED ALONG THEIR LENGTH (MARTÍNEz et al. 1996), AND IN UFRGS PV-1051-T THE DISTAL MARGIN bEARS A CONSPICUOUS SERRATION, WHEREAS, IN THE MESIAL bORDER, SERRATIONS ARE VISIbLE ONLY NEAR THE TIP Of THE TOOTH (OLIVEIRA et al. 2010). AS IN THE OTHER ECTENINIIDS, CAPPA / UFSM 0 0 29 SHOWS A SMALL DIASTEMA bETWEEN THE CANINE AND THE fIRST UPPER POSTCANINE ( FIGS. 2–3 View FIGURE 2 View FIGURE 3 ).
THERE ARE fIVE UPPER POSTCANINES MESIODISTALLY-ALIGNED IN CAPPA / UFSM 0 0 29 , PLACED IN THE LEfT MAXILLA. THEY ARE LITTLE EXPOSED, AND THEIR DENTAL ANATOMY IS POORLY PRESERVED ( FIG. 3 View FIGURE 3 ). THE fIRST ONE HAS ONLY ONE MAIN CUSP VISIbLE. HOWEVER THE DISTAL MARGIN Of THE TOOTH IS DAMAGED . BEHIND Of THE fIRST POSTCANINE, THERE IS A GAP LARGE ENOUGH TO ACCOMMODATE THREE TEETH (SECOND TO fOURTH POSTCANINES). THIS GAP VISIbLY PRESENTS A HEALED bONE, INDICATING NATURAL TEETH LOSS. THE SUPPOSED fIfTH, SIXTH, AND SEVENTH POSTCANINES ARE MORE EXPOSED AND COMPLETE THAN fIRST AND EIGHTH TEETH. THEY ARE LONGITUDINALLY ALIGNED, WITH TWO WELL-RECURVED AND POSTERIORLY DIRECTED CUSPS: ONE LARGE MESIAL MAIN CUP AND OTHER SMALLER ACCESSORY DISTAL ONE. WHEREAS THE NEW SPECIMEN EXPOSES ONLY THESE TWO CUPS, IN OTHER ECTENINIIDS THESE SAME ELEMENTS PRESENT THREE TO fOUR CUSPS. THE fIfTH TO SEVENTH POSTCANINES ARE SUbEqUAL IN SIzE. THE EIGHTH TOOTH IS SMALLER THAN THE fIfTH TO SEVENTH POSTCANINES. IT IS DAMAGED, PRECLUDING ACCESS TO ITS MORPHOLOGY.
THE LEfT SIDE Of THE SPECIMEN PVSJ 481 View Materials Of E. lunensis , WHICH IS AffECTED bY SIMILAR LATERAL COMPRESSION AS CAPPA / UFSM 0 0 29 , HAS A SIMILAR DENTAL MORPHOLOGY. THE LAST UPPER POSTCANINES, fIfTH TO SEVENTH, ARE MORE EXPOSED THAN THE OTHERS AND PRESENT TWO CUSPS, ONE PRINCIPAL AND LARGE, AND ANOTHER SMALL ACCESSORY. HOWEVER, IN THE RIGHT SIDE Of PVSJ 481 View Materials , WHICH IS NOT AffECTED bY COMPRESSION, THE TEETH HAVE THREE CUSPS PRESERVED.
AS IN OTHER ECTENINIIDS, THE POSTCANINES (fIfTH TO SEVENTH) Of CAPPA/UFSM 0 0 29 ARE SECTORIAL. IN CAPPA/ UFSM 0 0 29 AND OTHER ECTENINIIDS, ALL TEETH ARE IN CONTACT WITH EACH OTHER, fORMING AN IMbRICATED DENTITION WITH bACkWARDLY DIRECTED CUSPS ( FIG. 3 View FIGURE 3 ). IN T. riograndensis (UFRGS PV-1051-T) AND E. lunensis , THE fOUR ANTERIOR POSTCANINES ARE SMALLER THAN THE fOUR POSTERIOR ONES, THESE LAST PRESENTING MESIAL MAIN CUSPS LARGER THAN THE ACCESSORY ONES, WITH THEIR MESIAL MARGIN POSTERIORLY RECURVED (MARTÍNEz et al. 1996; OLIVEIRA et al. 2010). D. elegans PRESERVES ONLY THE fOURTH POSTCANINE, WHICH PRESENTS A COMPLETE CROWN WITH fOUR CUSPS (MARTÍNEz et al. 2013). THE RECURVED PATTERN Of THE MESIAL MARGIN RESEMbLES THE CONDITION fOUND IN OTHER ECTENINIIDS AND CHINIqUODONTIDS.
CAPPA / UFSM 0 0 29 LACkS SERRATIONS ON ANY Of ITS POSTCANINE TEETH . IN THE HOLOTYPE Of T. riograndensis , ALL THE CUSPS HAVE MESIAL AND DISTAL MARGINS SERRATED EXCEPT bY THE MESIAL EDGE Of THE MAIN CUSP Of THE fIVE fIRST POSTCANINES (OLIVEIRA et al. 2010). IN E. lunensis THE CUSPS Of ALL POSTCANINES ARE SERRATED IN bOTH EDGES (MARTÍNEz et al. 1996), AND IN D. elegans THE ONLY PRESERVED POSTCANINE (fOURTH), LACkS SERRATIONS (MARTÍNEz et al. 2013).
AS THE MANDIbLE IS IN THE POSITION Of ARTICULATION WITH THE SkULL , THE LOWER DENTITION Of CAPPA/UFSM 0 0 29 IS HIDDEN bY THE UPPER CHEEk TEETH SERIES AND THE MAXILLA, WITH ONLY A SMALL ANTERIOR PORTION Of THE LEfT CANINE VISIbLE. USING COMPUTED TOMOGRAPHY TO ACCESS THE INTERNAL ANATOMY Of THE SPECIMEN ( FIG. 8 View FIGURE 8 ), IT WAS POSSIbLE TO IDENTIfY THE PRESENCE Of THREE LOWER INCISORS AND AT LEAST SEVEN POSTCANINES. THE LOWER CANINES ARE ALSO ObSERVAbLE INSIDE THE PARACANINE fOSSA WITH THEIR TIPS SLIGHTLY PROTRUDING THROUGH THE SkULL ROOf; HOWEVER, THEY ARE NOT VISIbLE ON THE DORSAL SURfACE. REGARDING THE LOWER TEETH Of COMPARATIVE SPECIMENS, IN UFRGS PV-1051-T THERE ARE 3I /1C/8PC, AND IN E. lunensis (PVSJ 422) THE LOWER DENTITION IS UNkNOWN bECAUSE THE LOWER jAW IS WHOLLY OCCLUDED (MARTÍNEz et al. 1996; OLIVEIRA et al. 2010).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |