
Thinodromus dasys Gildenkov, 2000
|
publication ID |
https://doi.org/10.5281/zenodo.5739643 |
|
DOI |
https://doi.org/10.5281/zenodo.5911267 |
|
persistent identifier |
https://treatment.plazi.org/id/03CEDF09-FFF7-F345-D1AF-9089D80BFEA1 |
|
treatment provided by |
Marcus |
|
scientific name |
Thinodromus dasys Gildenkov, 2000 |
| status |
|
Thinodromus dasys Gildenkov, 2000 View in CoL
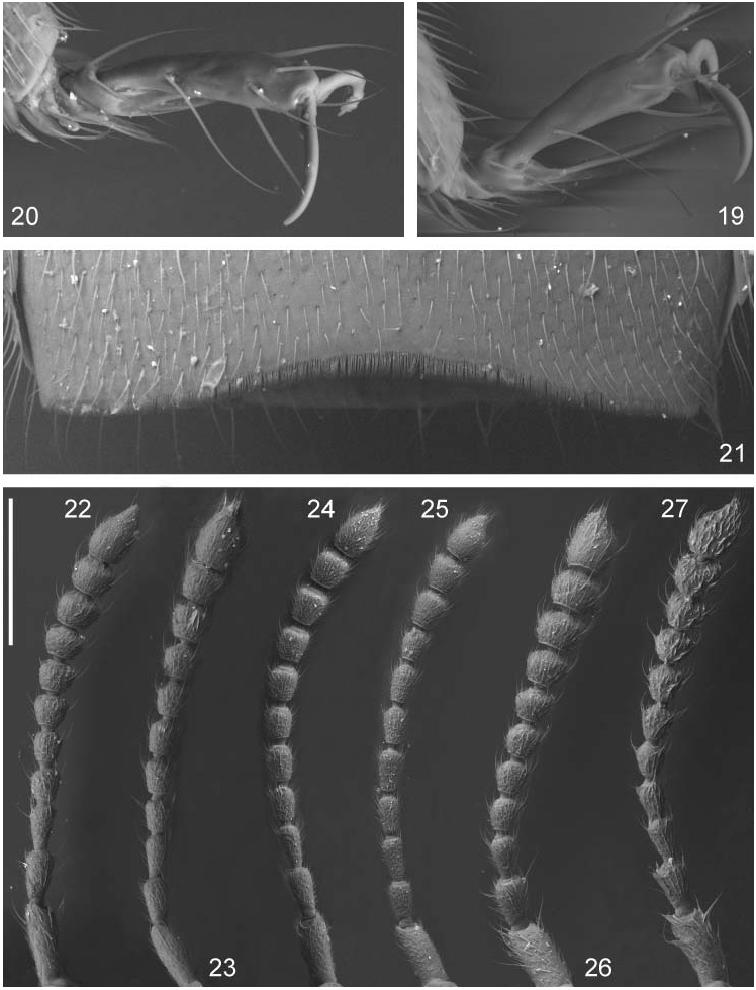
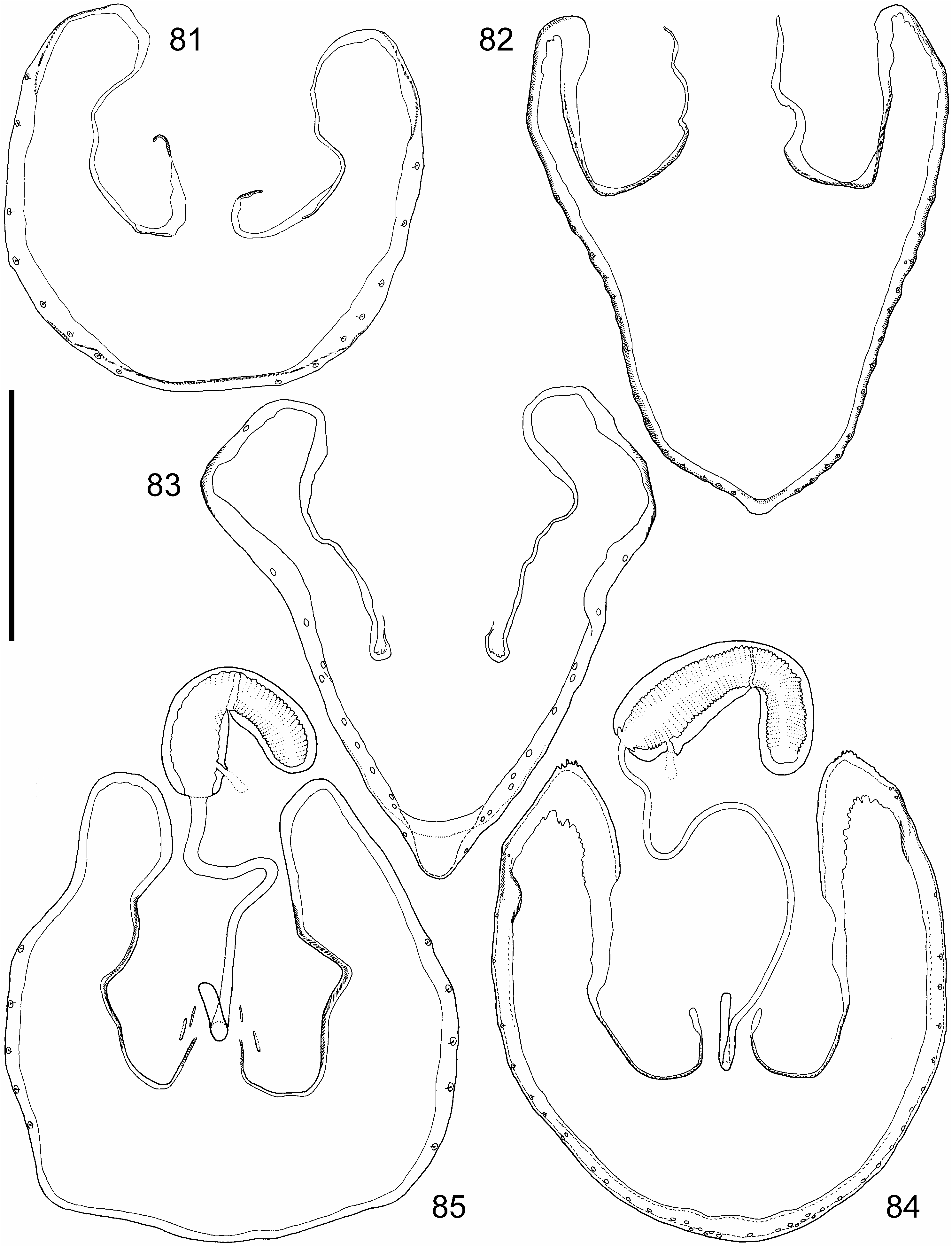
( Figs 5–6 View Figs 4–6. 4 , 26–27 View Figs 19–27. 19 , 40–42 View Figs , 46–47 View Figs 46–51. 46–47 , 82 View Figs 81–85. 81–83 )
Thinodromus ( Thinodromus) dasys Gildenkov, 2000: 51 View in CoL .
Thinodromus dasys: MAKRANCZY (2009: 38) View in CoL .
Material examined. REPUBLIC OF SOUTH AFRICA: N Transvaal [ Limpopo prov. = north part of former Transvaal], Woodside, Letsitele [ 23°53′S, 30°24′E]; 17.iii.1976, leg. R.E.Parrott (2 ♂♂ 1♀, CNCI; 1♂, MGSC), GoogleMaps same but 15.iii.1976 (1 ♂, CNCI); GoogleMaps N Transvaal, Kruger N.P., Satara [rest camp] [ 24°23′30″S, 31°46′30″E, 270m]; 7.iii.1976, leg. R.E. Parrott (1 ♂, CNCI) GoogleMaps ; N Transvaal, Kruger N.P., Skukuza research camp, 24°59′S, 31°36′E, 25.ii.1995, leg. S. Endrődy-Younga (E-Y 3120), UV light & trap (1♂ 1♀, TMSA, 1♂, ZMHB), GoogleMaps same but 25°00′S, 31°35′E, 19.ii.1995 (E-Y 3102) (4 ♂♂ 1 ♀, TMSA; 1 ♂, SMNS; 1 ♀, ZMHB), GoogleMaps same but 24°59′S, 31°35′E, 22.i.1995, leg. C.L. Bellamy (E-Y 3090) (3 ♂♂ 2 ♀♀ TMSA; 1 ♀, SMNS; 1 ♂ 1 ♀, NHMW), GoogleMaps same but 7.iii.1996, leg. S. Endrődy-Younga (E-Y 3220), UV light (1 ♂ 1 ♀, TMSA), GoogleMaps same but 1.iii.1995 (E-Y 3123), UV light & trap (2 ♂♂ 1 ♀, TMSA), GoogleMaps same but 3.iii.1996 (E-Y 3208), UV light (1 ♂, TMSA) GoogleMaps ; Natal, Hluhluwe N.P., 28°02′S, 32°05′E, 4–6.ii.1994, leg. M. Uhlig, lux ( 1 ♀, ZMHB) GoogleMaps . ZIMBABWE (former Rhodesia): Atlantica , 16mi W Salisbury [= Harare] [ 17°52′S, 30°49′E, 1370m], i.1974, leg. M.B. Fenton (1 ♂, CNCI). GoogleMaps
Redescription. Measurements (in mm, n = 10): HW = 0.63 (0.59–0.65); TW = 0.57 (0.54– 0.58); PW = 0.73 (0.68–0.76); SW = 0.85 (0.81–0.90); AW = 0.97 (0.90–1.03); HL = 0.40 (0.38–0.42); EL = 0.25 (0.23–0.26); TL = 0.033 (0.03–0.035); PL = 0.53 (0.49–0.55); SL = 0.84 (0.79–0.87); SC = 0.79 (0.72–0.82); FB = 1.82 (1.65–1.88); BL = 3.50 (2.81–3.98). Lustre and colour: Body rather shining but covered with pubescence on some parts not appearing so lustrous. Forebody almost pitch black, even antennae, mouthparts and legs (including tarsi) blackish dark brown. Abdomen also blackish dark brown but apices of tergites (where more transparent) slightly reddish. Shape and sculpture: Head ( Fig. 5 View Figs 4–6. 4 ) rather transverse, eyes prominent and occupy most of sides. Temples short but conspicuous, marked by a more or less discernible smooth rim around posterior edge of eyes; this rim forms with the rest of temple a moderately sharp edge. Pronotum transverse but slightly less so than in majority of species, first half of sides and anterior corners broadly rounded, side margin slightly concave in hind half, posterior angles rounded but not so obtuse, therefore discernible. Horseshoeshaped impression deep but not too broad, runs into a pair of depressions at sides of disc, two smaller impressions in middle of disc. Deflexed margin rather wide, conspicuous laterally and present at hind margin all along. Elytra ( Fig. 6 View Figs 4–6. 4 ) combined somewhat broader than long, slightly dilated posteriorly. Posterior elytral margin with a slightly pulled out membranous lobe near outer corner. Abdomen seems to be more constricted at base than usual for the genus and more parallel-sided. Apex of tergite VII with palisade fringe (widest medially). Punctation and microsculpture: Head punctation deep with punctures of quite uneven sizes and interspaces, latter occupies about half of the surface and free of microsculpture, rather shiny. Pronotum with similar punctation, some scabrous microsculpture in posterior corner. Elytra with even larger and deeper punctures, yet with ample shiny interspaces. Abdominal terga very roughly but sparsely punctured, puncture sizes decrease towards apical margin. Almost indiscernible traces of fine coriaceous microsculpture in the transversal groove posteriad basal ridges on tergites. Pubescence: Forebody with setae of varying sizes, rather sparse and mostly dark but predominantly long, erect setae; short interommatidial setae present without longer setae. Abdomen with sparse but long setae, with a few exceptionally long ones at apices of tergites. Finer and shorter setae only on head, pronotal disc and abdominal tergum bases. Primary and secondary sexual features: Female antennae ( Fig. 26 View Figs 19–27. 19 ) rather transverse, middle antennomeres (articles 4–5) about as long as broad, penultimate antennomeres (articles 9–10) about half longer than broad. Male antennae ( Fig. 27 View Figs 19–27. 19 ) with middle antennomeres (articles 4–5) distinctly elongate, penultimate antennomeres (articles 9–10) slightly broader than long. Male: MA of aedeagal internal sac elongate, somewhat spoon-like widened at apex, ML very transverse, BM extremely elongate, broadening medially, BA moderately long, distally widening, AC reverse V-shaped with arms forming an acute angle ( Figs 40–42 View Figs ), sternite VIII ( Fig. 46 View Figs 46–51. 46–47 ), tergite X ( Fig. 47 View Figs 46–51. 46–47 ); female: ringstructure ( Fig. 82 View Figs 81–85. 81–83 ).
Differential diagnosis. Of the here treated species, T. dasys is closely allied to T. gildenkovi sp. nov., although their external appearances differ greatly. Thinodromus dasys is also remarkably similar to T. decorsei (Cameron, 1948) , this species appears to be its sibling, supported by the slightly pointed but not widened tip and the widened, lamellar basal loops of the female ringstructure (a bit more elongate in T. dasys ), the sclerotization pattern of the basal part of the parameres, similar size and shape of the AC but in T. decorsei with an apical projection, MA elongate with spoon-like apex in both species, BM also very elongate, BM almost identical in shape, the medial tooth adjacent to ML is similar but slightly more bulky in T. dasys . The subapical edge of the median lobe body (on frontal side between paramere bases) is quite similar in both species, but the central sclerotized plate has a bifurcate apex in T. dasys .
Distribution. This species is known from the eastern Republic of South Africa and Zimbabwe.
Remarks. The original description was based on a single male specimen and now both sexes are known. This species was also referred to in MAKRANCZY (2009) as the closest known relative of Thinodromus decorsei (Cameron, 1948) giving the present treatment an extra value from a phylogenetic point of view. The specimen from Natal is without elytra.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Oxytelinae |
|
Genus |
Thinodromus dasys Gildenkov, 2000
| Makranczy, György 2013 |
Thinodromus ( Thinodromus ) dasys
| MAKRANCZY GY 2009: ) |
| GILDENKOV M. & YU 2000: 51 |