
Teratohyla sornozai, Cisneros-Heredia, Diego F., Yánez-Muñoz, Mario H. & Ortega-Andrade, Mauricio, 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.190232 |
|
DOI |
https://doi.org/10.5281/zenodo.5618692 |
|
persistent identifier |
https://treatment.plazi.org/id/B44A5E46-2D22-4272-FF43-384AFE7BFDC8 |
|
treatment provided by |
Plazi |
|
scientific name |
Teratohyla sornozai |
| status |
sp. nov. |
Teratohyla sornozai View in CoL sp. nov.
Figures 1–2 View FIGURE 1 View FIGURE 2
Holotype. DHMECN 3522, an adult female collected at a small stream tributary of Río Naranjal (00°21’05” N, 78°55’01” W, 750 m elevation), cantón Cotacachi, provincia de Imbabura, República del Ecuador, 0 1 May 2006 by H. M. Ortega-Andrade.
Paratypes. DHMECN 2634, adult male, Reserva Biológica Canandé (00°18’22.5” N, 79°08’17.6” W, 550 m elevation), cantón Quinindé, provincia de Esmeraldas, República del Ecuador, 21 August 2004, M. Yánez-Muñoz, P. Meza-Ramos, M. M. Reyes & A. Loaiza. DHMECN 4308, adult female, Bosque Protector Mashpi (00°10’2.34” N, 78°52’2.32” W, 1100 m), provincia de Pichincha, República del Ecuador, May 2007, M. H. Yánez-Muñoz y C. Castro.
Referred material. DHMECN 3521, a juvenile female with the same data of the holotype.
Diagnosis. A glassfrog species ( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 , 3 View FIGURE 3 E) diagnosed from all other Centrolenidae by the combination of the following characters: (1) vomerine teeth present; (2) snout truncate in dorsal view, protruding in profile; nostrils slightly elevated producing an slight depression in the internarial area; loreal region concave; (3) tympanic annulus evident, oriented dorsolaterally; very weak supratympanic fold above the tympanum; (4) dorsal skin slightly shagreen with minute granulations; (5) ventral skin coarsely areolate; subcloacal tubercles absent but subcloacal area granular; other cloacal ornamentation absent; (6) parietal peritoneum (condition P3), visceral peritoneum translucent except for pericardium covered by iridophores (condition V1); (7) liver tri-lobed (condition H0); (8) humeral crista dorsalis not forming a humeral spine in adult males; (9) webbing basal between fingers I and II; webbing formula for outer fingers II 1 −–3 III 2 −– 1IV ( Fig. 2 View FIGURE 2 ); (10) webbing on feet I 1 −–1 II 1 −–1− III 1 −–1− IV 1– 1 V ( Fig. 2 View FIGURE 2 ); (11) no dermal folds or tubercles on hands, upper arm or feet; ulnar and tarsal folds present (12) unpigmented nuptial pad Type I; concealed prepollex; (13) first finger longer than second, (14) eye diameter larger than width of disc on finger III; (15) colour in life, uniform bright green dorsum, green bones; (16) colour in preservative, dorsal surfaces deep grey-purple with few blurred cream spots; (17) iris golden copper with dark fine reticulations; (18) melanophores present on fingers and toes; (19) males call from upper side of leaves and ferns around smalls streams; call undescribed; (20, 21, 22) fighting behaviour, egg clutches, and tadpoles unknown; (23) snout-vent length in adult females 20.4–21.3 mm (n = 2); adult male paratype 19.0 mm.
Comparisons. Teratohyla sornozai ( Fig. 1 View FIGURE 1 ) is the only Glassfrog with protruding snout in profile, uniform green dorsal colouration, extensive webbing between outer fingers, and fully webbed toes. Extensive webbing between the outer fingers, humeral crista ventralis not developed into a humeral spine and uniform dorsal colouration also occurs in the following species: Rulyrana adiazeta (Ruiz-Carranza & Lynch) , Teratohyla amelie (Cisneros-Heredia & Meza-Ramos) , Rulyrana orejuela (Duellman & Burrowes) , T. spinosa (Taylor) , R. susatamai (Ruiz-Carranza & Lynch) , and Cochranella xanthocheridia Ruiz-Carranza & Lynch. However , all differ from T. sornozai as follows: Rurylana adiazeta and R. susatamai have rounded to truncate snouts in lateral profile, less extensive webbing between the fingers and not fully webbed toes, abundant melanophores on hands and feet, a different iris colouration (brown with dark brown reticulations), and they inhabit the eastern slopes of Cordillera Oriental and Central of Colombia. Teratohyla amelie has a rounded snout in lateral profile, iridophores covering the visceral peritonea, transparent parietal peritoneum, and inhabits the central and southern lowlands and foothills of Amazonian Ecuador. Rulyrana orejuela has a truncate snout in lateral profile, less webbing between the outer fingers and not fully webbed toes, abundant melanophores on hands, feet, and ventral surfaces, different iris colouration (dark grey with yellow ring around pupil), and larger SVL in adult males (27.3–28.3 mm). Teratohyla spinosa has a truncate snout in lateral profile, less webbing between the outer fingers and not fully webbed toes, and adult males have a prepollical spine at the base of the thumb. Cochranella xanthocheridia has a rounded to truncate snout, less webbing between the outer fingers and not fully webbed toes, and it inhabits the northern portion of the Cordillera Occidental of Colombia. Further, in preservative, T. sornozai shows a deep grey-purple colouration, while all other species show a lavender colouration.
Description of the holotype. Adult female, SVL = 21.3 mm (Fig. 20). Body slender. Head distinct, slightly wider than long, and wider than body; HW/HL = 1.05, HW/SVL = 0.37, HL/SVL = 0.35. Snout short, truncate in dorsal view, protruding in profile, EN/HL = 0.30; nostrils slightly elevated producing a slight depression in the internarial area; canthus rostralis indistinct; concave loreal region; lips flared. Eyes large, ED/HL = 0.35, directed anterolaterally at about 45° from midline, eyes can be seen when viewed from below, interorbital area wider than eye diameter, IOD/ED = 1.54, EN/ED = 0.87, EN/IOD = 0.56. Tympanic annulus evident, oriented dorsolaterally with dorsal inclination; weak supratympanic fold from behind the eye to the insertion of the arm, separated from orbit by a longer distance than the tympanum diameter. Dentigerous processes of vomers present, choanae moderately sized, rounded, near margin of mouth; tongue rounded, slightly notched behind, not indented posteriorly.
Skin of dorsal surfaces of head, body, and limbs slightly shagreened with minute granulations. Ventral surfaces coarsely areolate; cloacal opening directed posteriorly at upper level of thighs; no distinct cloacal sheath, some small low tubercles around the cloacal opening; subcloacal tubercles absent, other cloacal ornamentation absent.
Upper arm thin, forearm moderately robust, breadth of upper arm about half that of forearm. Humeral spine absent; ulnar fold present, tubercles absent. Relative lengths of fingers III> IV> I> II; webbing basal between fingers I and II, webbing formula for other fingers I 1–3 III 2 −– 1 IV; bulla absent; finger discs slightly wide, truncate; disc on third finger slightly larger than those on toes, and shorter than eye diameter, 3DW/ED = 0.62; all subarticular tubercles rounded and elevated; indistinct supernumerary tubercles present in fingers; palmar tubercle large, rounded, plane; thenar tubercle elliptic, rather indistinct.
Hind limbs slender; TL/SVL = 0.63, FL/SVL = 0.53. Tarsal fold present; inner metatarsal tubercle large and elliptical, flat; outer metatarsal tubercle indistinct. Subarticular tubercles rounded and low; supernumerary tubercles small, rather indistinct. Webbing on foot I 1 −–1 II 1 −–1− III 1 −– 1− IV 1– 1 V; discs truncate on all toes, slightly expanded.
Measurements of the holotype. Snout-vent length, 21.3 mm; head width, 7.9 mm; head length, 7.5 mm; horizontal eye diameter, 2.6 mm; inter-orbital distance, 4.0 mm; eye-nostril distance, 2.3 mm; internarial distance between the nostrils, 1.8 mm; width of disc on the third finger, 1.6 mm; tibia length, 13.3 mm; foot length, 11.2 mm.
Colouration. In life ( Fig. 1 View FIGURE 1 , 3 View FIGURE 3 E), dorsal surface uniform bright green, yellowish towards the flanks, without light or dark flecks, spots, or ocelli. Upper eyelids yellowish-green or bright green; tips of digits and interdigital webbing dull yellow to greenish-yellow. Throat and ventral surfaces of legs bluish-green. Parietal peritoneum white (condition P3). Bones pale green. Iris golden copper to golden brown with dark thin reticulations.
In preservative, dorsum deep grey-lavender with few blurred cream spots; dorsal surfaces of arms greylavender, hind limbs cream-greenish or lavender; upper eyelid dark blue; lips cream. All ventral surfaces cream. Parietal peritoneum white (condition P3); all visceral peritonea clear except for the white pericardium (condition V1).
Specimen DHMECN 3522 DHMECN 4308 DHMECN 2634 Variation. Variation of measurements and body proportions is presented in Tables 1 View TABLE 1 and 2 View TABLE 2 . Photographs of specimens in life are shown in Figs. 1 View FIGURE 1 and 3 View FIGURE 3 . Gross morphological and colouration features are fairly invariant; male paratype, adult female paratype, and juvenile female are similar to female holotype. Hand and foot webbing does not vary ( Fig. 2 View FIGURE 2 ). In preservative a few blurred cream irregular spots appear in some specimens (DHMECN 3521–22), while other remain uniform lavender. These spaces are produced by the absence of melanophores rather than by the presence of light pigmentation (iridophores or xanthophores). Humeral spine and prepollical spine absent in both sexes. Adult male is smaller than adult females (SVL male =19.0 mm; SVL females = 20.4–21.3 mm). Males have unpigmented nuptial pads Type –I.
Specimen DHMECN 3522 DHMECN 4308 DHMECN 2634 Etymology. The specific name of this Glassfrog is a noun in the genitive case and a patronym for our friend Francisco (Pancho) Sornoza, Ecuadorian ornithologist and conservationist. He is president of Fundación Jocotoco, an Ecuadorian NGO that manages the Reserva Biológica Canandé, and has actively support the research activities of the Museo Ecuatoriano de Ciencias Naturales that resulted in the discovery of this new species.
Distribution and natural history. Teratohyla sornozai is known from three localities in the provinces of Esmeraldas, Imbabura, and Pichincha, northwestern Ecuador ( Fig. 4 View FIGURE 4 ). This species occur in Foothill Evergreen Forests (DHMECN 3521, 3522, 4308) and Lowland Evergreen Forests (DHMECN 2634), with an altitudinal range between 550 and 1100 m elevation. The female holotype and juvenile female were found sitting on leaves of herbs and ferns up to 2 meters above a small stream, Naranjal River. No egg clutches or tadpoles were seen in the area. At Naranjal, T. sornozai was found in sympatry with Espadarana prosoblepon (Boettger) , Pristimantis labiosus (Lynch, Ruiz-Carranza & Ardila-Robayo) , and Hyloscirtus alytolylax (Duellman) . Although just females where found at Naranjal, males were calling in the surroundings but over 5 meters above the stream. At Canandé, the male was found calling from the upper surface of a leave and in sympatry with E. prosoblepon , Sachatamia albomaculata (Taylor) , S. ilex (Savage) , Hyloscirtus palmeri (Boulenger) , Hypsiboas boans (Linnaeus) , and H. picturatus (Boulenger) . At Mashpi, the male was collected in sympatry with Pristimantis labiosus (Lynch, Ruiz-Carranza & Ardila-Robayo) , E. prosoblepon , Hyloscirtus palmeri , and H. picturatus .
Remarks. Teratohyla sornozai is similar to T. spinosa , sharing a similar body form and size, uniform dorsal colouration, lobed liver, ventral parietal peritoneum covered by iridophores, and gastrointestinal and hepatic peritonea without iridophores. We hypothesise that both are sister taxa based on their high morphological similarity, but this remains to be tested. Teratohyla sornozai has not been found in sympatry with T. spinosa . It is possible that both occur together particularly on lowland areas. In contrast with T. spinosa that prefers low riverine vegetation along relatively quiet areas (Kubicki, 2007; pers. obs.); T. sornozai apparently prefers higher strata along riverine vegetation (over 1.5 m) especially in the splash zone of waterfalls.
We have not found egg clutches of Teratohyla sornozai but we have observed egg clutches of T. amelie , T. midas , T. pulverata , and T. spinosa . Females of T. amelie , T. midas , and T. pulverata deposit their egg clutches on the upper side of leaves, while T. spinosa may use both sides of leaves. Eggs are laid in a laminar array ( Altig and McDiarmid 2007) at different locations on the leave (centre or borders, showing intra-specific variation). The egg clutches of T. amelie , T. midas and T. spinosa keep the original arrangement and location where they were deposited, but the egg clutches of T. pulverata frequently absorb water and swell considerably, forming a 3-dimensional clump that moves due to gravity to the tip of the leave and hangs ( Cisneros-Heredia & McDiarmid 2007: 28). Eggs of all four species have a greenish-tinted colouration (Kubicki 2007; pers. obs.), which is apparently produced by the accumulation of biliverdin in the eggs, and seems to be common to all members of this clade, probably a synapomorphy.
Teratohyla sornozai corresponds to the species mentioned as Cochranella sp. N 11 by Yánez-Muñoz & Cisneros-Heredia (2008).

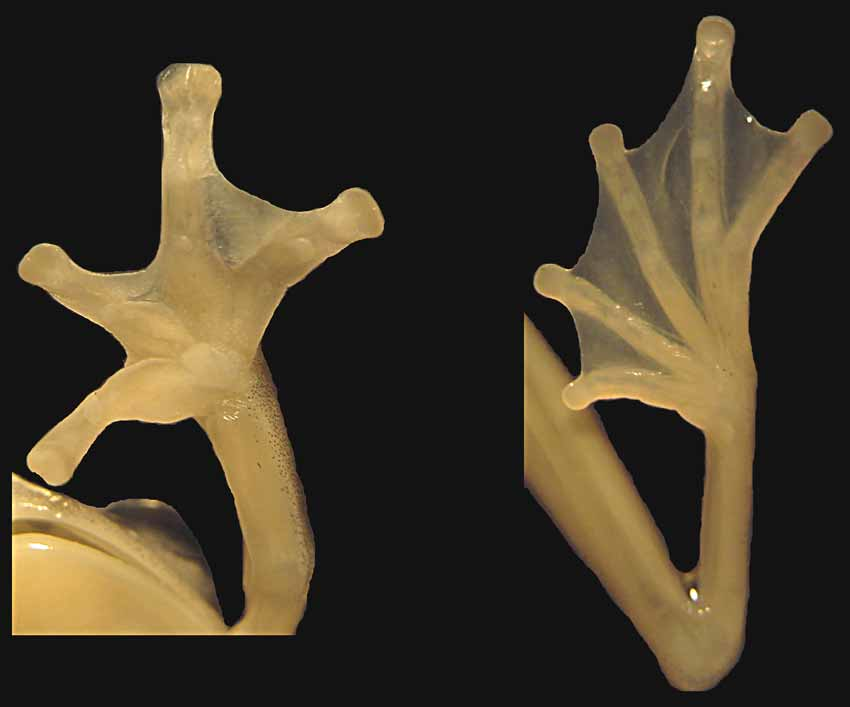

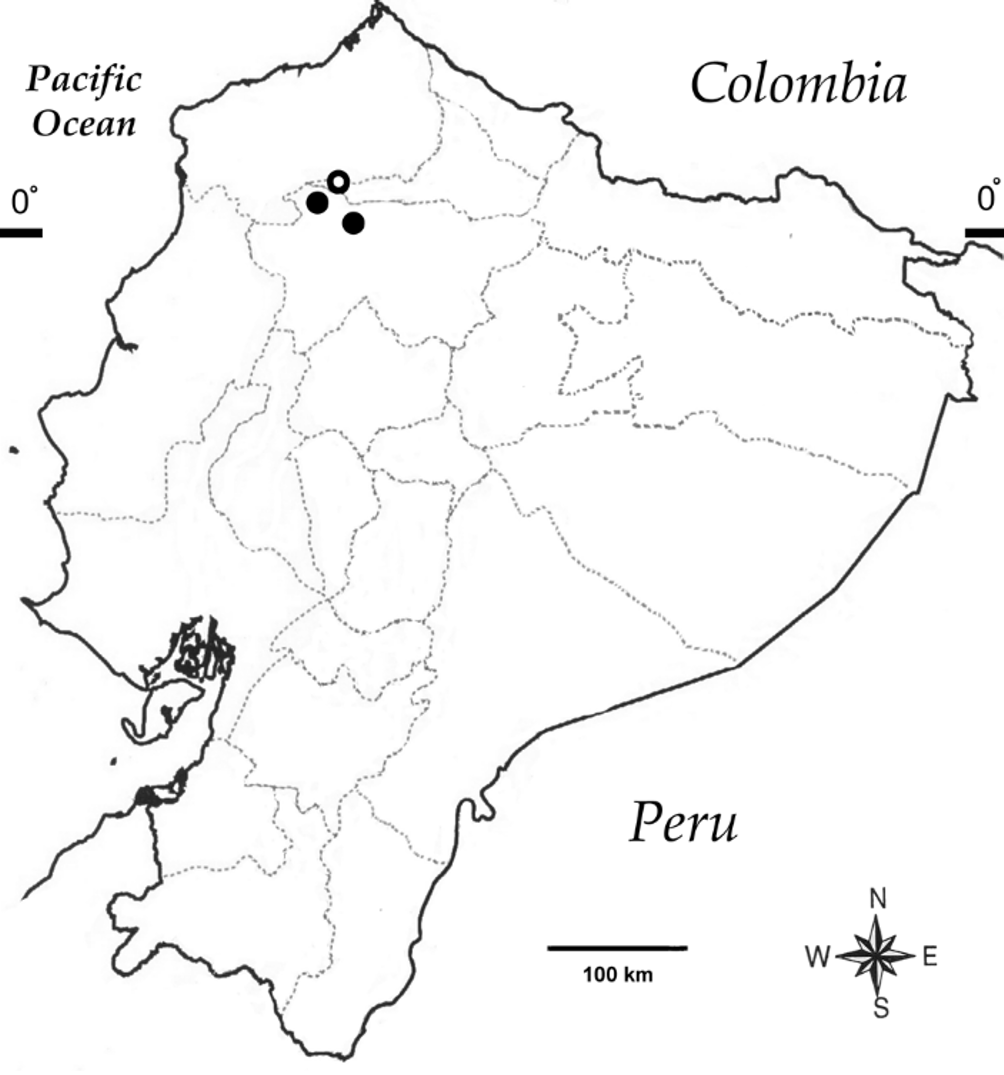
TABLE 1. Variation of measurements (in mm) of adult Teratohyla sornozai sp. nov. See text for abbreviations.
| Holotype | Paratype | Paratype | |
|---|---|---|---|
| Sex | Female | Female | Male |
| SVL | 21.3 | 20.4 | 19.0 |
| HW | 7.9 | 7.3 | 6.5 |
| HL | 7.5 | 7.2 | 6.7 |
| ED | 2.6 | 2.8 | 2.3 |
| IOD | 4.0 | 2.6 | 3.5 |
| EN | 2.3 | 2.5 | 2.4 |
| IN | 1.8 | 1.7 | 1.7 |
| TD | 0.9 | 0.9 | 0.8 |
| 3DW | 1.6 | 1.6 | 1.2 |
| TL | 13.3 | 13.7 | 11.2 |
| FL | 11.2 | 13.3 | 8.8 |
TABLE 2. Variation of proportions of adult Teratohyla sornozai sp. nov. See text for abbreviations.
| Holotype | Paratype | Paratype | |
|---|---|---|---|
| Sex | Female | Female | Male |
| HW/HL | 1.01 | 1.05 | 0.97 |
| HW/SVL | 0.36 | 0.37 | 0.34 |
| HL/SVL | 0.35 | 0.35 | 0.35 |
| EN/HL | 0.34 | 0.30 | 0.35 |
| EN/HW | 0.34 | 0.29 | 0.36 |
| EN/ED | 0.88 | 0.87 | 1.02 |
| EN/IOD | 0.95 | 0.56 | 0.67 |
| ED/HL | 0.39 | 0.35 | 0.35 |
| ED/HW | 0.38 | 0.33 | 0.36 |
| ED/3DW | 1.79 | 1.63 | 1.92 |
| IOD/ED | 0.93 | 1.54 | 1.52 |
| IOD/HW | 0.36 | 0.51 | 0.54 |
| IOD/HL | 0.36 | 0.53 | 0.53 |
| IN/IOD | 0.66 | 0.44 | 0.49 |
| TD/ED | 0.32 | 0.33 | 0.35 |
| TL/SVL | 0.67 | 0.63 | 0.59 |
| FL/SVL | 0.65 | 0.53 | 0.46 |
| TL/FL | 1.03 | 1.19 | 1.27 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |