
Spiralothelphusa wuellerstorfi ( Heller, 1862 )
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.798.1629 |
|
publication LSID |
lsid:zoobank.org:pub:AF8AAD44-164C-4514-BACF-16961F3EAABF |
|
DOI |
https://doi.org/10.5281/zenodo.6328979 |
|
persistent identifier |
https://treatment.plazi.org/id/B2722D05-1323-1F39-FF72-7DDAC75D8426 |
|
treatment provided by |
Felipe |
|
scientific name |
Spiralothelphusa wuellerstorfi ( Heller, 1862 ) |
| status |
|
Spiralothelphusa wuellerstorfi ( Heller, 1862) View in CoL
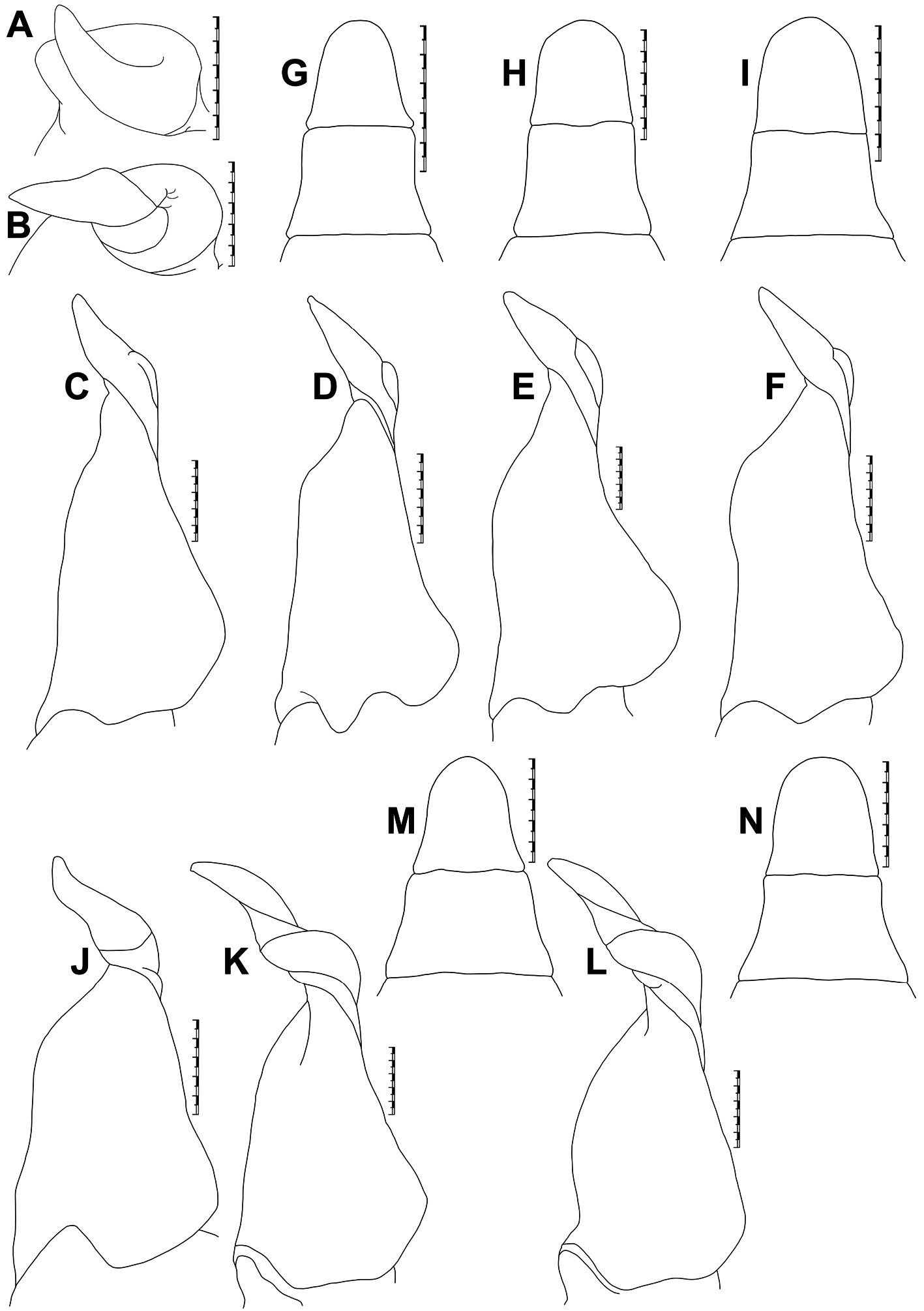
Figs 3–6 View Fig View Fig View Fig View Fig , 7B, K, M View Fig
Thelphusa wüllerstorfi Heller, 1862: 520 .
Paratelphusa innominata Fernando, 1960: 210 , figs 6f, 12b, 13c, d, 14, 15 (part).
Thelphusa leschenaudii – Heller 1865: 32 (not Thelpheusa leschenaudii H. Milne Edwards, 1837 View in CoL ).
Telphusa leschenaultii – Wood-Mason 1871: 202 (part) (not Thelpheusa leschenaudii H. Milne Edwards, 1837 View in CoL ).
Telphusa leschenaulti – Henderson 1893: 382 (part). — Ortmann 1893: 489. — Bürger 1894: 2 (not Thelpheusa leschenaudii H. Milne Edwards, 1837 View in CoL ).
Potamon aurantium – Ortmann 1897: 302 (part) (not Cancer aurantius Herbst, 1799).
Potamon wüllerstorfi – De Man 1898: 438 (list).
Potamon (Potamon) hydrodromus – Rathbun 1904: 287, not pl. 12 fig. 3 (part) (not Cancer hydrodroma Herbst, 1794 View in CoL ).
Spiralothelphusa wuellerstorfi View in CoL – Bott 1968: 404, figs 3–4, 26; 1970: 97, pl. 18 figs 12–14, pl. 29 fig. 68. — Ng & Tay 2001: 190, figs 23a–k, 24a–b, h–l, 25a, b (part). — Beenaerts et al. 2010: 188, 190, tables S1, S4 (listed only in phylogenetic tree and supplementary files). — Pati & Sudha Devi 2015: 416 (part).
Diagnosis
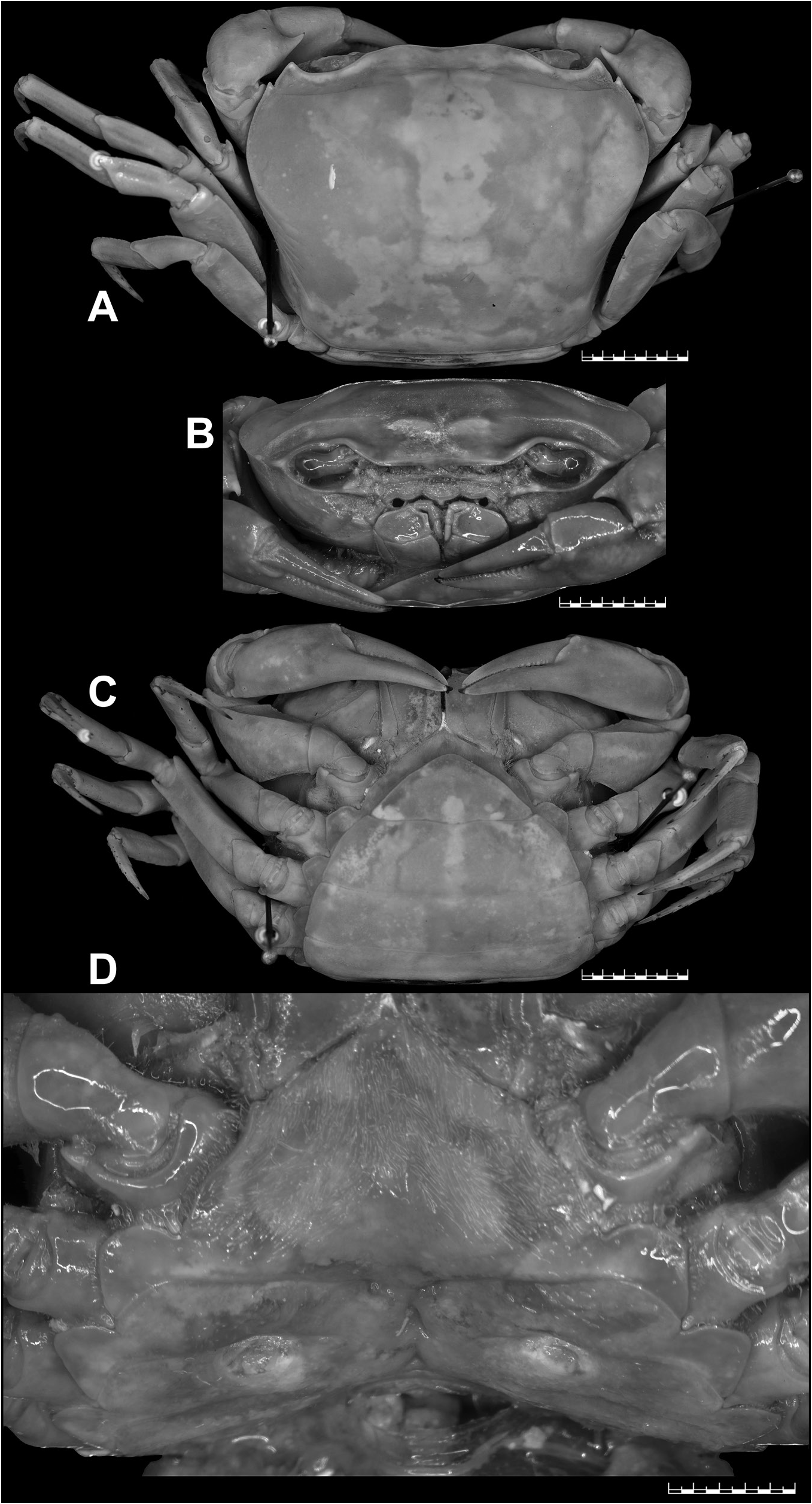
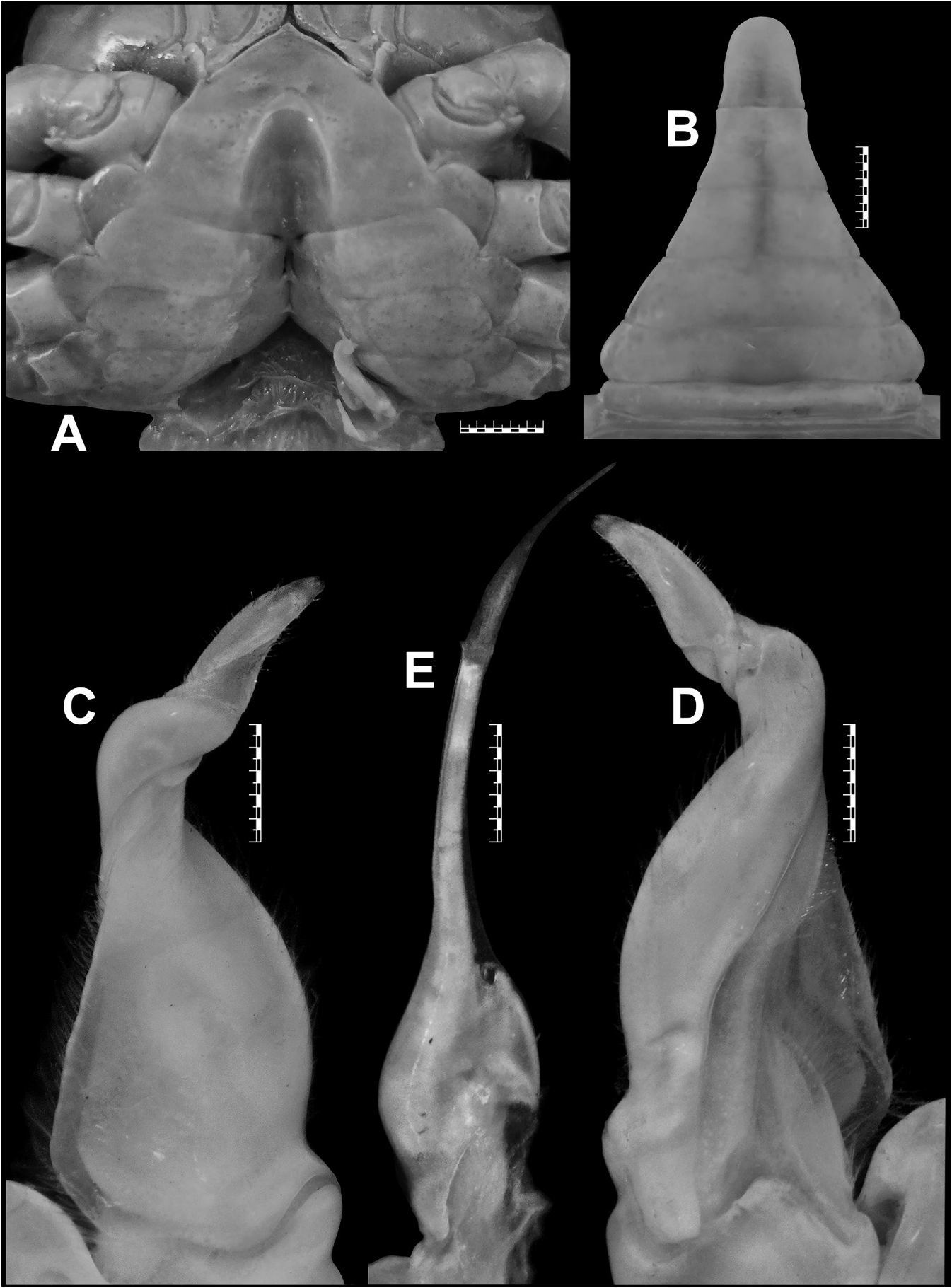
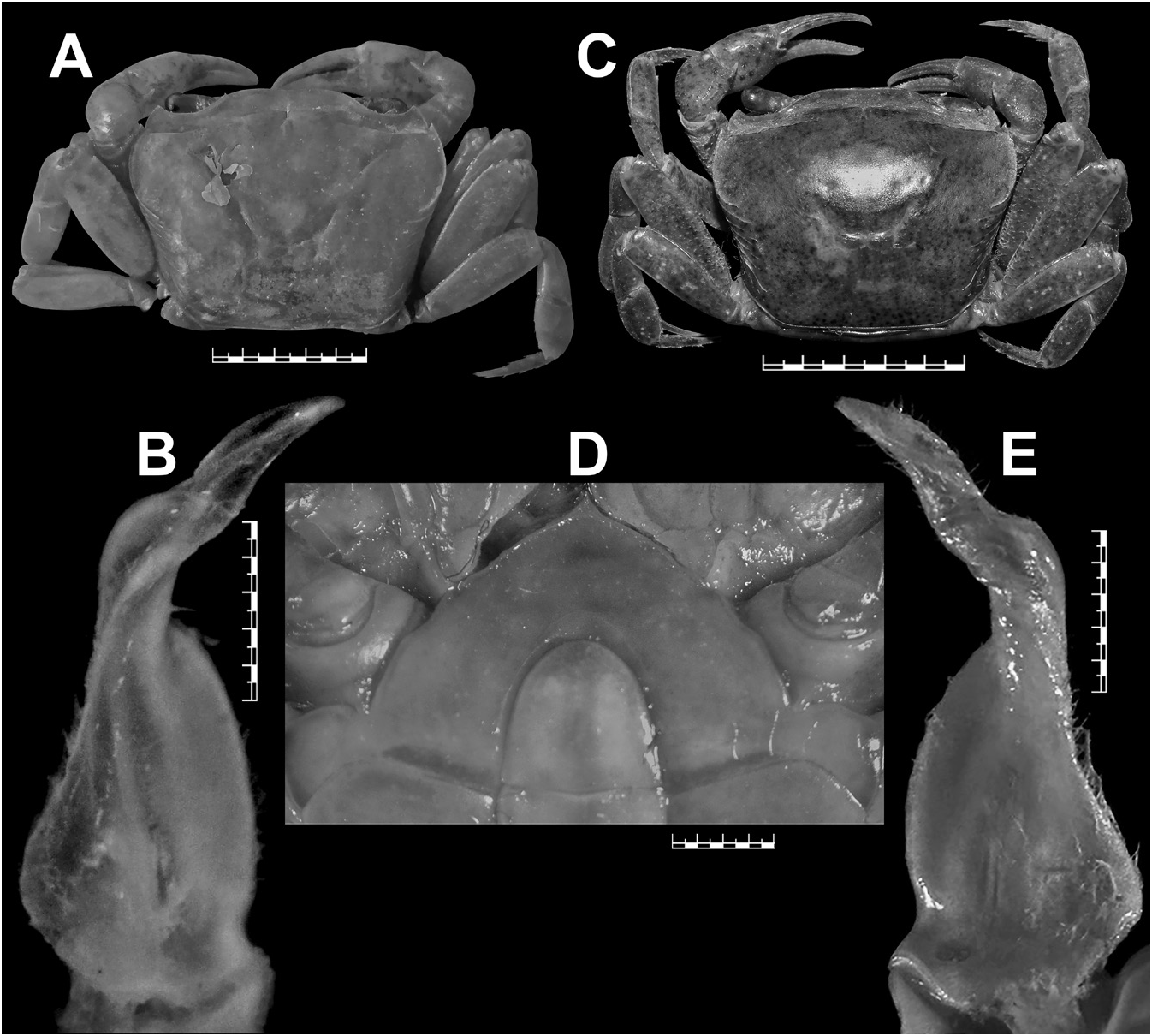
Carapace broader than long (CW/CL=1.3), deep ( CH /CL=0.7–0.8); dorsal surface strongly convex in frontal view; epigastric cristae well-developed, sharp, distinctly anterior to postorbital cristae; postorbital cristae sharp, distinctly curved; external orbital angle triangular; epibranchial tooth distinct, blunt to slightly sharp, above level of postorbital cristae; branchial regions gently inflated; cervical grooves shallow, narrow, discontinuous, not reaching to level of postorbital cristae; suborbital margin concave, discontinuous with supraorbital margin; frontal median triangle complete, as broad as frontal margin, lateral margins joining dorsal margin; frontal margin medially concave in dorsal view; epistomal medial lobe with distinct, blunt tooth ( Figs 3A–B View Fig , 5A–B View Fig ). Eyes relatively small as compared to orbital space ( Figs 3B View Fig , 5B View Fig ). Third maxilliped exopod with well-developed flagellum. Chelipeds smooth, subequal or unequal; chela with elongated dactylus and palm; carpus with long, narrow, sharp inner distal major tooth and very low sub-basal granule; merus lacking subterminal spine ( Figs 3A–C View Fig , 5A–C View Fig ). Ambulatory legs moderately stout, shorter than chelipeds ( Figs 3A, C View Fig , 5A, C View Fig ). Male S1 and S2 separated by deep groove; male S2/S3 visible as shallow medial depression, not reaching lateral margins; male S3/S4 only visible as shallow lateral depressions ( Figs 3C View Fig , 4A View Fig ). Male sternopleonal cavity reaching to imaginary line joining anterior part of cheliped coxae ( Figs 3C View Fig , 4A View Fig ). Male pleon narrowly triangular, with concave lateral margins; somite 6 trapezoidal, relatively broader (proximal width circa 1.6 times medial length), with gently concave lateral margins ( Figs 3C View Fig , 7M View Fig ). Male telson slightly longer than pleonal somite 6, with concave lateral margins and narrow apex ( Figs 3C View Fig , 7M View Fig ). G1 relatively slender, with tip gently hooked downwards; terminal segment and distal portion of subterminal segment longitudinally strongly twisted full turn, clockwise in left G1 and anticlockwise in right G1; twisted portion long, circa 0.5 times length of non-twisted portion; non-twisted portion with outer margin sharply, relatively less strongly curved, straight towards base ( Figs 4B–C View Fig , 6A–C View Fig , 7K View Fig ). G2 circa 1.0–1.1 times length of G1; distal segment long, circa 0.5 times length of basal segment ( Fig. 4D View Fig ). Female pleonal somites and telson in adult broadly ovate ( Fig. 5C View Fig ). Vulvae in adult on S6 positioned apart from each other (VD/SW=circa 0.3), each ventrally opened, subovate, large (occupying circa 0.6 times length of S6), positioned close to S5/S6 ( Fig. 5D View Fig ).
Material examined
Lectotype (designated herein) INDIA • 1♂, adult(CW 37.4 mm, CL 28.5mm,CH 19.6 mm, FW 12.7 mm);“Nicobaren”[sic];1857–1859; Novara exped.; NHMW CR 27063 View Materials (formerly NHMW CR 24937 View Materials , labelled as ‘ Thelphusa Leschenaudii M. Edw., Nicobaren, Frfld. ’).
Paralectotypes INDIA • 1 ♀, adult (CW 37.8 mm, CL 28.6 mm, CH 21.6 mm, FW 12.4 mm) ; same collection data as for lectotype; NHMW CR 27064 View Materials (formerly NHMW CR 24937 View Materials , labelled as ‘ Thelphusa Leschenaudii M. Edw., Nicobaren, Frfld. ’) • 4 ♂♂ (32.3 × 24.5 mm; 31.2 × 24.4 mm; 30.1 × 23.5 mm; 26.8 × 21.3 mm), 4 ♀♀ (23.9 × 18.0 mm; 23.8 × 19.5 mm; 23.7 × 18.8 mm; 23.6 × 18.6 mm); same collection data as for lectotype; NHMW CR 1613 View Materials • 2 ♂♂ (35.8 × 27.9 mm, gonopods removed; 28.2 × 22.6 mm); same collection data as for lectotype; NHMW CR 24936 View Materials • 3 ♀♀ (37.8 × 29.2 mm; 34.5 × 27.5 mm; 32.3 × 25.3 mm); same collection data as for lectotype; NHMW CR 24937 View Materials • 9 subadults (26.0 × 20.2 mm; 25.6 × 18.9 mm; 20.6 × 16.1 mm; 18.8 × 14.6 mm; 18.8 × 14.4 mm; 18.6 × 13.9 mm; 17.6 × 13.2 mm; 17.3 × 12.8 mm; 15.4 × 11.6 mm); same collection data as for lectotype; NHMW CR 24938 View Materials .
SRI LANKA • 1 ♂ (28.0 × 23.0 mm, gonopods removed), 1 ♀ (28.2 × 21.5 mm), 3 subadults (15.1 × 11.2 mm; 13.8 × 10.7 mm; 12.3 × 10.1 mm); “Ceylon” [sic]; 1857–1859; Novara exped.; NHMW CR 24939 View Materials .
FRENCH POLYNESIA • 2 ♂♂ (36.3 × 28.4 mm; 33.0 × 23.9 mm); “Taiti” [sic]; 1857–1859; Novara exped.; NHMW CR 24940 View Materials • 4 ♂♂ (32.5 × 25.1 mm, right gonopods removed; 32.3 × 24.4 mm; 31.3 × 23.5 mm, right gonopods removed; 30.5 × 23.6 mm), 3 ♀♀ (36.7 × 28.3 mm; 35.9 × 28.2 mm; 34.9 × 27.3 mm); “ Taiti ” [sic]; 1857–1859; Novara exped.; NHMW CR 24941 View Materials • 3 ♂♂ (36.5 × 27.3 mm; 35.2 × 27.3 mm; 34.1 × 26.0 mm), 2 ♀♀ (37.1 × 29.0 mm; 33.9 × 27.1 mm); “Taiti” [sic]; 1857–1859; Novara exped.; NHMW CR 24942 View Materials .
Additional material
INDIA – Tamil Nadu State • 3 ♂♂, 1 ♀; Chennai District, Puzhal Lake, near Pammathukulam; 13.159° N, 80.159° E; alt. 15 m; 19 Sep. 2020; H. Sruthi leg.; ZSI- WRC C.1995 GoogleMaps • 9 ♂♂, 5 ♀♀; Kanchipuram District, Valathottam, near Kanchipuram; 12.784° N, 79.704° E; alt. 79 m; 16 Nov. 2019; P. Kalaimagal et al. leg.; ZSI-WRC C.1960 GoogleMaps • 1 ♂; Kanchipuram District, paddy fields in front of Madras Crocodile Bank, Mamallapuram [= Mahabalipuram ]; 12.745° N, 80.220° E; alt. 1 m; 7 Dec. 1997; M.M. Bahir leg.; ZRC 2003.0243 View Materials (formerly WHT 10749 ) GoogleMaps .
Description of male lectotype
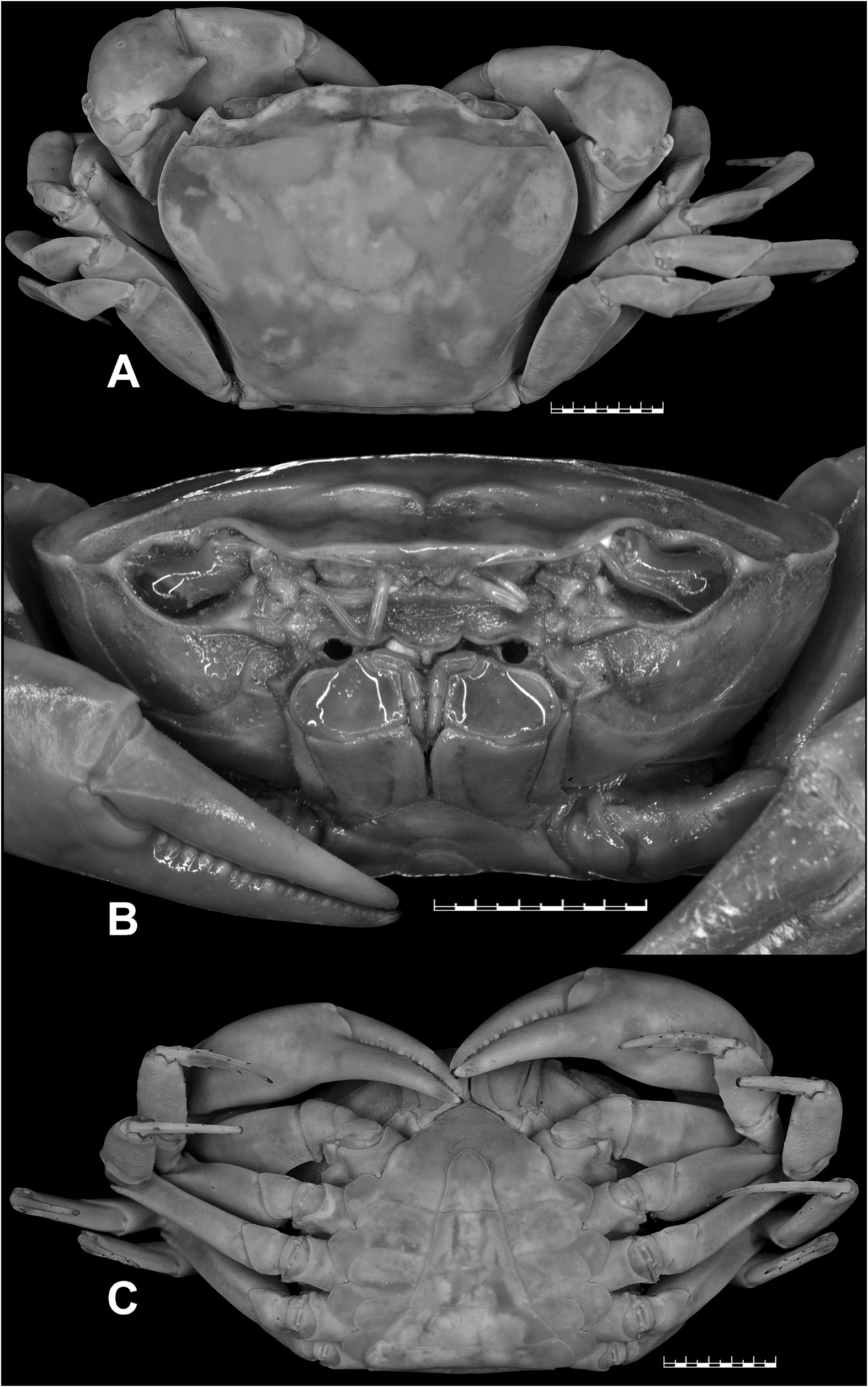
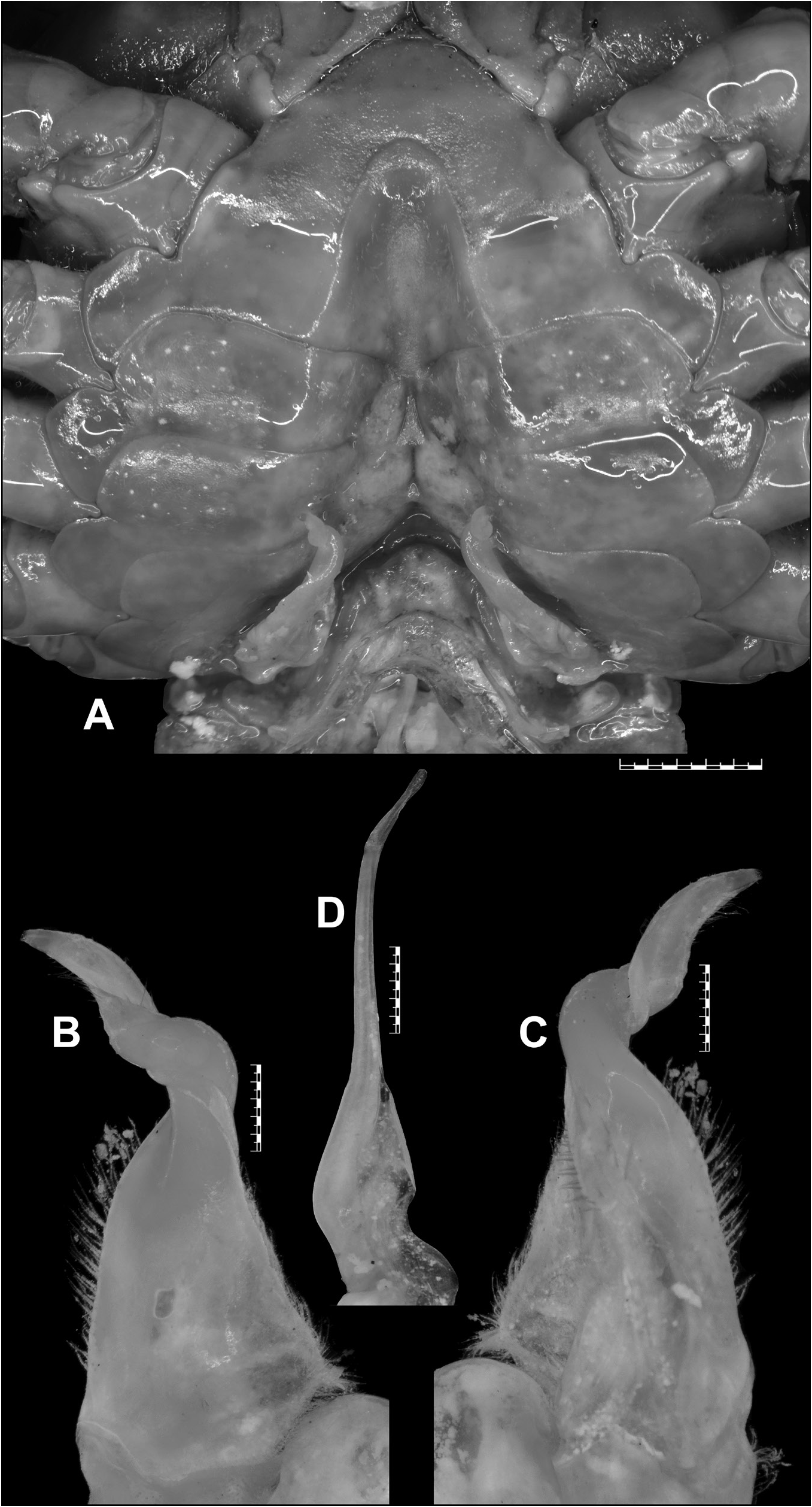
Carpace broader than long (CW/CL=1.3), deep ( CH /CL= 0.7); dorsal surface strongly convex in frontal view, smooth except for epigastric cristae, postorbital cristae and posterolateral surfaces; anterolateral margins gently convex, cristate, shorter than posterolateral margins; posterolateral margins medially concave; epigastric cristae well-developed, sharp, distinctly anterior to postorbital cristae; postorbital cristae sharp, distinctly curved, each sloping posteriorly towards postorbital region, then anteriorly towards anterolateral margin; external orbital angle triangular, with outer margin circa 2 times length of inner margin; epibranchial tooth distinct, blunt, above level of postorbital cristae; postorbital region concave; branchial regions gently inflated; cervical grooves shallow, narrow, discontinuous, not reaching to level of postorbital cristae; mesogastric groove deep, narrow, short, bifurcated posteriorly; H-shaped groove distinct; subhepatic region rugose; suborbital region almost smooth; pterygostomial region smooth; supraorbital margin cristate, smooth, medially straight; suborbital margin concave, cristate with low granules, discontinuous with supraorbital margin; frontal median triangle complete, as broad as frontal margin, lateral margins joining dorsal margin; front gently sloping downwards, subtrapezoidal; frontal margin smooth, cristate, broad (FW/CW= 0.3), medially concave in dorsal view; epistomal medial lobe with distinct, blunt tooth ( Fig. 3A–B View Fig ).
Eyes relatively small as compared to orbital space; eyestalk short, narrow, laterally concave; cornea moderately large, pigmented ( Fig. 3B View Fig ).
Antennules long, folded in longitudinally broad fossae; antennae very long, reaching beyond eyestalk ( Fig. 3B View Fig ). Mandibular palp 2-segmented; terminal segment bilobed. First, second maxillipeds each with long flagellum on exopod. Third maxillipeds cover most of buccal cavity when closed; ischium subrectangular, longer than broad, with deep, oblique submedial groove; merus subrectangular, broader than long, sunken; exopod moderately stout, distally narrow, longer than ischium, reaching proximal half of merus, with well-developed flagellum ( Fig. 3B View Fig ).
Chelipeds smooth, unequal, left chela larger. Major chela with blunt teeth on each finger, very small gape when fingers closed; dactylus curved, slender, elongated; palm smooth, elongated; carpus generally smooth, gently inflated, with long, narrow, blunt (eroded) inner distal major tooth and very low subbasal granule; merus generally smooth, lacking subterminal spine ( Fig. 3A–C View Fig ).
Ambulatory legs moderately stout, shorter than chelipeds, P3 longest, glabrous; merus (P2–P5) lacking subdistal spine; dactylus (P2–P5) gently recurved, subequal in length to propodus, with few distinct, sharp chitinous spines on margins ( Fig. 3A, C View Fig ).
Thoracic sternites smooth, glabrous, sparsely punctate; S1 and S2 separated by deep groove; S2/S3 visible as shallow medial depression, not reaching lateral margins; S3/S4 only visible as shallow lateral depressions; S4/S5, S5/S6, S6/S7 deep, narrow, medially interrupted; S7/S8 shallow, narrow, with medial transverse ridge, interrupting longitudinal groove ( Figs 3C View Fig , 4A View Fig ). Pleonal locking mechanism with prominent tubercle on submedial part of S5 ( Fig. 4A View Fig ). Sternopleonal cavity deep, long, reaching to imaginary line joining anterior part of cheliped coxae ( Figs 3C View Fig , 4A View Fig ).
Pleon narrowly triangular, with concave lateral margins; somites 1, 2 narrower than somite 3; somite 3 broadest, with strongly convex lateral margins; somites 4, 5 trapezoidal, with straight lateral margins; somite 6 trapezoidal, broader than long (proximal width circa 1.6 times medial length), distinctly longer than preceding somites, with gently concave lateral margins ( Figs 3C View Fig , 7M View Fig ). Telson slightly longer than pleonal somite 6, slightly longer than broad (medial length circa 1.1 times proximal width), with concave lateral margins and narrow apex ( Figs 3C View Fig , 7M View Fig ).
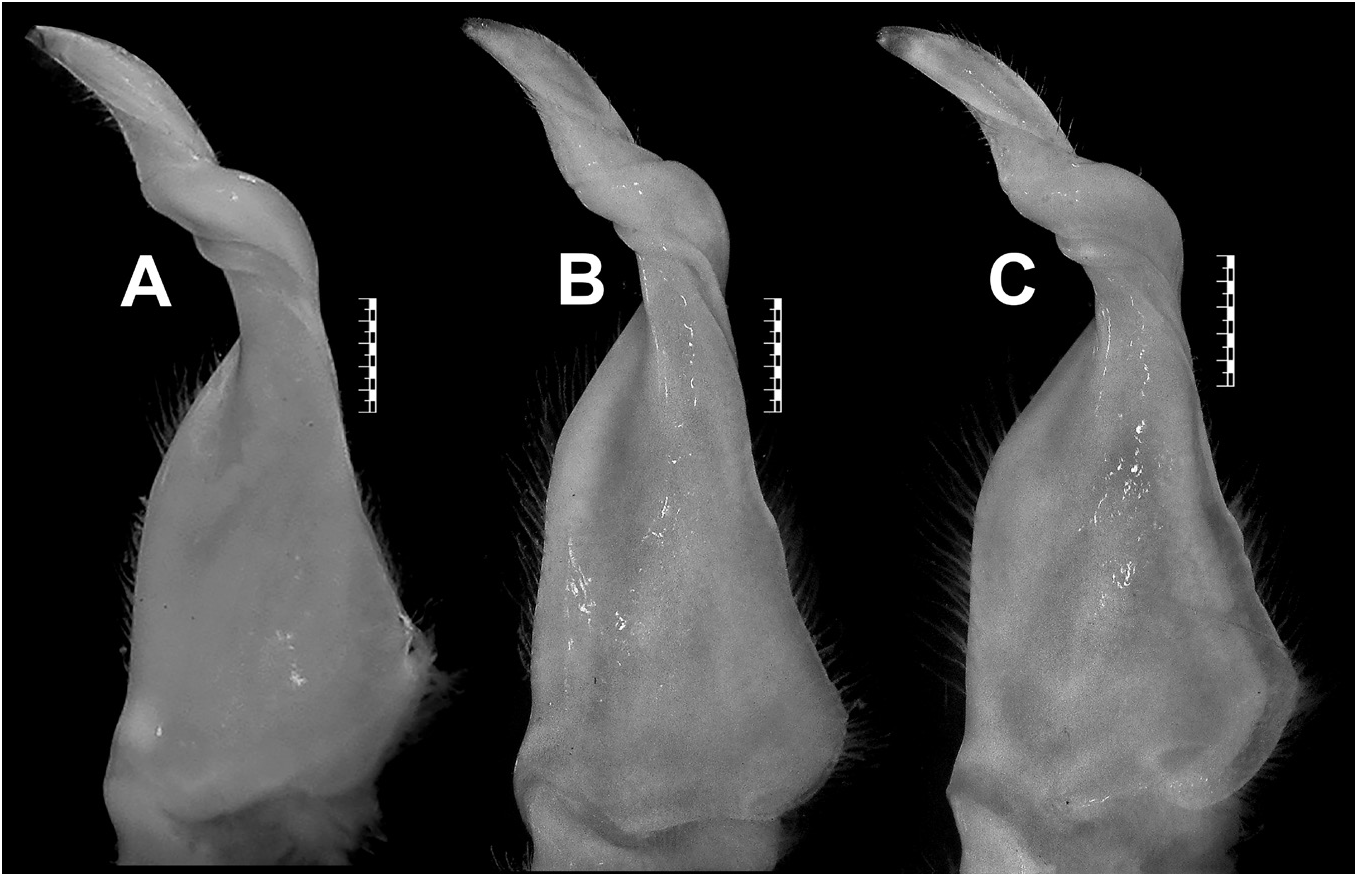
G1 relatively slender, with tip gently hooked downwards, not reaching S5/S6; terminal segment and distal portion of subterminal segment longitudinally strongly twisted full turn, clockwise in left G1 and anticlockwise in right G1; twisted portion long, circa 0.5 times length of non-twisted portion; non-twisted portion with outer margin sharply curved, straight towards base; groove for G2 lateral ( Figs 4A–C View Fig , 7K View Fig ). G2 circa 1.1 times length of G1; distal segment long, circa 0.5 times length of basal segment ( Fig. 4D View Fig ).
Remarks
Bott (1968) listed the lectotype (male) and seven paralectotypes (four males and three females) for S. wuellerstorfi from NHMW, with locality as “Taiti”, most probably the Nicobar Islands. Neither the specific catalogue number nor the size of the lectotype was mentioned by Bott (1968). Bott also never illustrated the lectotype the lectotype male designated by him. Instead, he provided the figures of a male specimen from “ Nikobaren ” located in the Forschungsinstitut Senckenberg, Frankfurt am Main, Germany (SMF 4406) (see Bott 1968: figs 3–4, 26; 1970: pl. 18 figs 12–14, pl. 29 fig. 68). NHMW possesses three lots (NHMW CR 24940–24942) from “Taiti”, and the lectotype is supposedly designated from the lot NHMW CR 24941 that actually contains four males and three females. Peter Dworschak, the former curator of the Crustacea collections of NHMW, confirmed that the lectotype designated by Bott (1968) was not marked specifically among the paralectotypes, and the right gonopods have been removed from two males but kept in the same vial. Without accurate information on catalogue number and size of the specimen, and in the absence of any illustration, the particular lectotype is now difficult to trace among several paralectotypes of the depository, even from the most likely lot NHMW CR 24941 (Peter Dworschak, personal communication). Bott (1968), therefore, did not unambiguously select a particular syntype to act as the unique name-bearing type of S. wuellerstorfi as required by Article 74.5 of the International Code of the Zoological Nomenclature ( ICZN 1999). A new lectotype should be now designated to avoid further confusion and to stabilize the taxonomy of the species. NHMW possesses 43 syntypes (17 males, 14 females, and 12 juveniles). Among these syntypes, the largest male is from the lot NHMW CR 24937 that contains this lone male and four females. This largest and intact adult male (37.4 × 28.5 mm) (NHMW CR 24937) from “Nicobaren” is hereby designated as the lectotype, with a new catalogue number (NHMW CR 27063), for the nomenclatural stability of S. wuellerstorfi . The presently designated lectotype is described and illustrated ( Figs 3A–C View Fig , 4A–D View Fig , 7K, M View Fig ). One of the female paralectotypes from the lot NHMW CR 24937 (now with a new catalogue number, NHMW CR 27064) is also illustrated here ( Fig. 5A–D View Fig ).
The female paralectotype (NHMW CR 27064) of S. wuellerstorfi examined in this study is an adult and possesses most of the non-sexual character states ( Fig. 5A–C View Fig ) as those in the male lectotype designated herein. Its epibranchial tooth, however, is relatively sharper ( Fig. 5A View Fig ) than that of the male lectotype ( Fig. 3A View Fig ). The female paralectotype (NHMW CR 27064) has almost equal-sized chelipeds, each with a sharp inner distal major tooth on the carpus ( Fig. 5A–C View Fig ), whereas the male lectotype possesses unequal chelipeds, each with an eroded inner distal major tooth on the carpus that appears blunt ( Fig. 3A–C View Fig ). The pleonal somites and telson of the female paralectotype (NHMW CR 27064) are broadly ovate, which cover the thoracic sternum except for the lateral edges when closed. The pleonal somite 1 is the shortest; pleonal somites 2–5 are progressively longer; pleonal somite 6 is the longest, much broader than long, slightly longer than the telson, with the convex lateral margins ( Fig. 5C View Fig ). The telson is triangular, much broader than long, with almost straight lateral margins ( Fig. 5C View Fig ). The vulvae on S6 of the female paralectotype (NHMW CR 27064) are positioned apart from each other (VD/SW = circa 0.3), each opens ventrally, subovate, large (occupying circa 0.6 times the length of S6), positioned close to S5/S6, and surrounded by a raised, broad rim ( Fig. 5D View Fig ).
The paratype male (MNHN BP 183) of S. wuellerstorfi from Nicobar illustrated in Ng & Tay (2001: fig. 24h–l), which should now be regarded as the paralectotype, is identical with the lectotype male (NHMW CR 27063) in the G1 structure. The male specimen from Sri Lanka (MNHN BP 11437; formerly labelled as ‘ Spiralothelphusa hydrodroma ’; probably determined by Rathbun) also has a similar G1 (see Ng & Tay, 2001: 24 a–b) as in the lectotype male (NHMW CR 27063) of S. wuellerstorfi ( Figs 4B View Fig , 7K View Fig ). Ng & Tay (2001), however, doubted the locality data of this MNHN specimen from Sri Lanka.
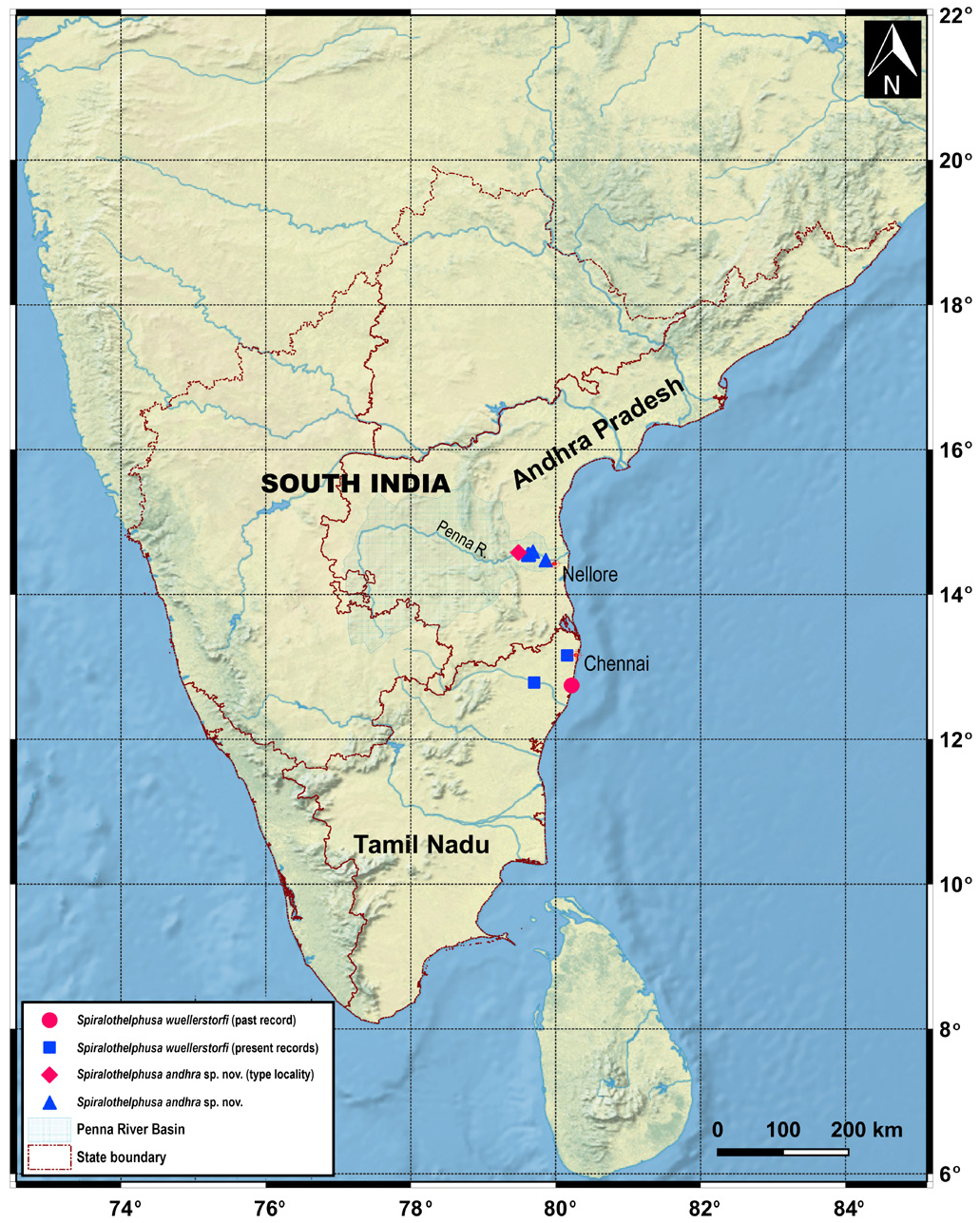
Spiralothelphusa wuellerstorfi was originally collected during the Novara expedition between 1857– 1859 from four distant localities, viz., Ceylon [= present-day Sri Lanka], Nicobaren [= present-day Nicobar Islands of India], Madras [= present-day Chennai of Tamil Nadu State], and Taiti [= Tahiti of French Polynesia] ( Heller 1862). The locality labels from the Novara expedition were obviously wrong (Peter Dworschak, personal communication). All the species of Spiralothelphusa are restricted to mainland India and Sri Lanka ( Ng & Tay 2001; Pati & Sudha Devi 2015). In fact, no primary freshwater crab belonging to Gecarcinucidae is so far known from the Nicobar Islands of India (cf. Pati & Thackeray 2018). While documenting the brachyuran crabs of the Andaman and Nicobar Islands, Dev Roy & Nandi (2012) commented that there was no report of S. wuellerstorfi since Heller (1862, 1865). A recent expedition (during 2018) to the Nicobar Islands by the Zoological Survey of India also could not locate S. wuellerstorfi or any other gecarcinucid species (K.A. Subramanian, personal communication). The records of S. wuellerstorfi from Nicobar and Tahiti are thus incorrect. All the Sri Lankan species of freshwater crabs appear to be endemic (Bahir et al. 2005). Moreover, no species of Spiralothelphusa from Sri Lanka has a G1 like that of S. wuellerstorfi . Spiralothelphusa wuellerstorfi is, therefore, less likely to occur in Sri Lanka. It might be originated from “Madras”, or possibly somewhere in the Tamil Nadu State, near the present-day Chennai. Scherzer (1861) while narrating the Novara expedition mentioned that the tanks (or artificial basins) in the Vellore region of Tamil Nadu supply irrigation water for rice fields, which could be the preferred habitat of S. wuellerstorfi . Unfortunately, no type material of S. wuellerstorfi from “Madras” could be located in NHMW (Peter Dworschak, personal communication). It seems none of the original localities of S. wuellerstorfi is correct.
Other known localities of S. wuellerstorfi are the Malabar Coast [= present-day the northern part of the Kerala State in the south-western coast of India] (H. Milne Edwards 1853; A. Milne-Edwards 1869; Wood-Mason 1871; Henderson 1893; Ortmann 1893, 1897; Rathbun 1904; Ng & Tay 2001; Rajesh et al. 2017), the Mulugu District of the Telangana State ( Pati & Mandal 2021), and the Tadoba National Park of the Maharashtra State ( Pati & Sharma 2014 sensu Pati & Sudha Devi 2015). These localities are also incorrect since the G1s of the available specimens or illustrated specimens (see Pati & Sharma 2014: fig. 11d–e; Ng & Tay 2001: 24 c–f are completely different from the lectotype (NHMW CR 27063) designated herein for S. wuellerstorfi . In fact, these specimens do not have a resemblance with the known species of Spiralothelphusa in the G1 structure. Beenaerts et al. (2010) included a specimen (WHT 10793) of S. wuellerstorfi from Mamallapuram [= Mahabalipuram] of the Kanchipuram District of Tamil Nadu in their phylogenetic analysis. The specimen of S. wuellerstorfi mentioned by Beenaerts et al. (2010) could not be traced in the most likely depository (ZRC). An adult male (ZRC 2003.0243; formerly WHT 10749) from the same locality (see additional material section), however, possesses an identical G1 ( Fig. 6A View Fig ) as that of the lectotype male (NHMW CR 27063) of S. wuellerstorfi ( Figs 4B View Fig , 7K View Fig ). Furthermore, the G1 of the holotype male (NHM 1892.7.15.241) of Paratelphusa innominata Fernando, 1960 , from Madras illustrated in Ng & Tay (2001: fig. 23i) is similar to that of the lectotype male (NHMW CR 27063) of S. wuellerstorfi ( Figs 4B View Fig , 7K View Fig ). Ng & Tay (2001) argued that Paratelphusa innominata could be a junior synonym of S. wuellerstorfi , albeit some specimens of the former species from Sri Lanka reported by Fernando (1960) are now known as S. fernandoi ( Ng 1994; Ng & Tay 2001). Ng et al. (2008) also listed Paratelphusa innominata as one of the two primary synonyms of S. wuellerstorfi without any comment. We now confirm Paratelphusa innominata to be a junior subjective synonym of S. wuellerstorfi as their G1s have no difference. As such, the previous records of S. wuellerstorfi from Mamallapuram and “Madras” of Tamil Nadu are reliable. Some recently collected specimens of S. wuellerstorfi (see additional material section) from Chennai and Kanchipuram districts of Tamil Nadu further confirm the presence of S. wuellerstorfi in the state as those specimens resemble the lectotype male (NHMW CR 27063) in the G1 structure ( Figs 4B View Fig , 6B, C View Fig , 7K View Fig ).
Heller (1865) provided emended characters for Thelphusa leschenaudii H. Milne Edwards, 1837 , followed by a brief description, which are almost identical to the Latin description of Thelphusa wüllerstorfi in Heller (1862). Heller (1865) also mentioned the same localities for Thelphusa leschenaudii as in Heller (1862) for Thelphusa wüllerstorfi . Although Heller (1865) did not provide any figure or illustration for Thelphusa leschenaudii , the description and localities indicate the species to be Spiralothelphusa wuellerstorfi . In any case, Thelphusa leschenaudii is a junior objective synonym of Telphusa indica Latreille, 1825 , which itself is a junior synonym of Oziotelphusa aurantia (Herbst, 1799) (see Bahir & Yeo 2005).
The species reported in Wood-Mason (1871), Henderson (1893), Ortmann (1893, 1897), Bürger (1894), De Man (1898), and Rathbun (1904) includes S. wuellerstorfi as presently defined as well as other taxa, viz., an unidentified species of Spiralothelphusa , Oziotelphusa aurantia , Oziotelphusa ceylonensis ( Fernando, 1960) , and Oziotelphusa ganjamensis (Pati & Sharma, 2012) , as evident from the locality records and/or published sources ( Fernando 1960; Ng & Tay 2001; Bahir & Yeo 2005; Pati & Vargila 2019).
In addition to Paratelphusa innominata , Thelphusa corrugata Heller, 1865 , was included as a junior synonym of S. wuellerstorfi by Ng et al. (2008) without further explanation. Thelphusa corrugata was originally described from Madras and Java during the Novara expedition ( Heller 1865). According to Cumberlidge (1997), Thelphusa corrugata is a junior synonym of Potamonautes perlatus (H. Milne Edwards, 1837) of the family Potamonautidae Bott, 1970 . Since Potamonautes perlatus is an African species, the original localities of Thelphusa corrugata are certainly erroneous ( Cumberlidge 1997). In a recent communication, Peter Ng informed that he has no clue how and why Thelphusa corrugata was assigned to S. wuellerstorfi in Ng et al. (2008), but that was an error, he now confirms based on the comments from Cumberlidge (1997).
Spiralothelphusa wuellerstorfi possesses a G1 with strongly, longitudinally twisted terminal segment and distal portion of the subterminal segment (fully turned clockwise in the left G1 and anticlockwise in the right G1) like that in S. parvula and S. andhra sp. nov. ( Figs 4B–C View Fig , 6A–C View Fig , 7B, J–L View Fig , 10C–D View Fig , 11B, E View Fig ; see Ng & Tay 2001). Like S. andhra sp. nov., S. wuellerstorfi is nevertheless separated from S. parvula by the relatively slender G1, with the tip gently hooked downwards ( Figs 4B–C View Fig , 6A–C View Fig , 7K–L View Fig , 10C–D View Fig , 11B, E View Fig ) (vs relatively stouter G1, with the tip strongly curved upwards in S. parvula ; Fig. 7J View Fig ). On the other hand, S. wuellerstorfi can be distinguished from S. andhra sp. nov. in the shape of the male telson and G1 non-twisted portion (see Remarks for S. andhra sp. nov.). Spiralothelphusa wuellerstorfi need not be confused with the remaining Spiralothelphusa species as it possesses a G1 with fully turned terminal segment and distal portion of the subterminal segment ( Fig. 7B View Fig ) (vs G1 with half turned terminal segment and distal portion of the subterminal segment in S. fernandoi , S. gibberosa , S. hydrodroma , and S. senex ; Fig. 7A View Fig ; see Ng & Tay 2001).
Geographic distribution
Spiralothelphusa wuellerstorfi is precisely known only from a few localities in the northeast of the Tamil Nadu State, South India ( Fig. 8 View Fig ) (also see Remarks for the species).
Diagnosis
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Gecarcinucoidea |
|
Family |
|
|
Genus |
Spiralothelphusa wuellerstorfi ( Heller, 1862 )
| Pati, Sameer K., Mandal, Sudipta & Jaiswal, Deepa 2022 |
Paratelphusa innominata
| Fernando C. H. 1960: 210 |
wüllerstorfi
| De Man J. G. 1898: 438 |
Thelphusa leschenaudii
| Heller C. 1865: 32 |
Thelphusa wüllerstorfi
| Heller C. 1862: 520 |