
Diopsidae
|
publication ID |
https://doi.org/ 10.3853/j.0067-1975.63.2011.1585 |
|
persistent identifier |
https://treatment.plazi.org/id/03C1878D-A60D-917C-FEA9-FC335CA695BE |
|
treatment provided by |
Felipe |
|
scientific name |
Diopsidae |
| status |
|
The Diopsidae View in CoL View at ENA
The broad classification of the Diopsidae given by Hennig (1965: 62) is still preferred. This gives two subfamilies: Centrioncinae , including Centrioncus (synonym Teloglabrus , since added; D. McAlpine, 1997); and Diopsinae, including all other genera. I have examined details of antennal structure in Centrioncus decoronotus Feijen , Sphyracephala (Hexechopsis) beccarii (Rondani) , and Cyrtodiopsis sp. (West Malaysia). Feijen (1983: figs 4–6) has illustrated the general features of some diopsid antennae.
The antennae of Diopsidae differ from those of the four diopsoid families treated above in the following conditions: segment 2, though having the rim well developed and encircling the distal articular surface, is not produced into a pair of dorsal lobes; the conus is relatively large, well removed from the medial margin of the distal articular surface, and the armature of the annular ridge is relatively slightly developed; caesti are absent; segment 3 has a large basal hollow, into which the conus is inserted, and has no basal stem—two features sharply distinguishing it from that of all other diopsoid families, including the Syringogastridae .
In Centrioncus ( Fig. 122 View Figures 122–125 ) the conus is deep, somewhat bilaterally compressed, and asymmetrical, with laterally facing preapical foramen; segment 3 ( Fig. 124 View Figures 122–125 ) has its basal foramen inside the basal hollow on its lateral wall; the arista is inserted slightly laterally to the dorsal margin of segment 3.
In Sphyracephala segment 2 ( Fig. 123 View Figures 122–125 ) is more nearly radially symmetrical (only slightly bilaterally compressed), with the conus erect, elongate, slightly clavate, arising from near the centre of the distal articular surface, and possessing a terminal foramen; the button is located preapically on the mediodorsal part of the conus (a most unusual location in the Cyclorrhapha); segment 3 ( Fig. 125 View Figures 122–125 ) has the basal hollow deep, almost symmetrical, with the basal foramen located on its floor; the arista is placed symmetrically on its dorsal margin. In Cyrtodiopsis the conus is even longer than in Sphyracephala , with the button on the dorsal surface at c. the distal quarter of its length. The form of segment 2 and its conus in these more advanced diopsids is remarkably convergent with that of the lower cyclorrhaphous family Lonchopteridae .
Other genera of Diopsinae (e.g., Diopsis ) appear similar to the above examples in most details. The antenna of Centrioncus is more like that of various basal schizophoran types found in the Sciomyzoidea and Heteromyzoidea than is that of the Diopsinae, though it may partly retain the plesiomorphic structure from which that of the Diopsinae was derived.
The lower tephritoid families
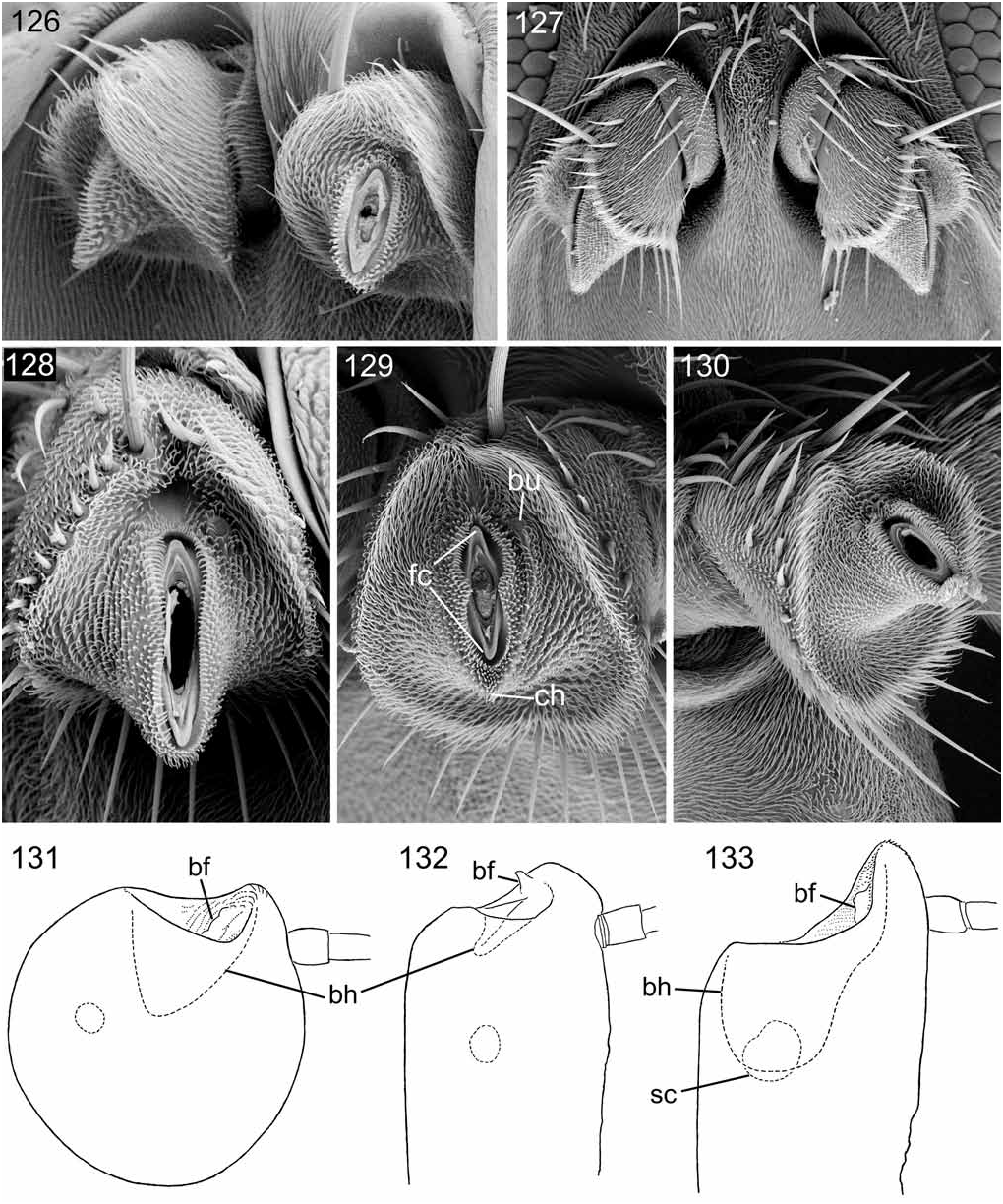
In this informal category I include the families Lonchaeidae , Pallopteridae , Piophilidae , Richardiidae , and Ulidiidae (syn. Otitidae ) as distinct from the higher tephritoid families Pyrgotidae , Platystomatidae , and Tephritidae . I have examined in at least moderate detail the antenna of one species of each family, viz. Lonchaea sp. (Lonchaeidae) , Palloptera muliebris (Harris) (Pallopteridae) , Piophila vitrea McAlpine (Piophilidae) , Richardia tephritina Enderlein (Richardiidae) , and Herina macalpinei Kameneva (Ulidiidae) ; see Figs 126–133 View Figures 126–133 .
Wiegmann et al. (2011) in their phylogenetic study of dipterous families remove the Pallopteridae far from the tephritoid families, with which they are usually associated, and place the family as the sister group of the Neurochaetidae . I find antennal segment 2 of Palloptera to be exceedingly similar structurally to that of the tephritoid genus Richardia Fig. 129 View Figures 126–133 ) and without the distinctive characters of the Neurochaetidae . This fact, together with the distinctive tephritoid synapomorphies of the pallopterid female postabdomen, not approached in the Neurochaetidae (see J. McAlpine, 1987: figs 5, 6; D. McAlpine, 1978: fig. 19), and the general lack of morphological and behavioural resemblances between the two families render the placement indicated by Wiegmann et al. very improbable. Also, the plesiomorphic Eocene neurochaetid genus Anthoclusia shows no significant points of resemblance to the Pallopteridae ( Hennig, 1965, and author’s unpublished study).
The lower tephritoid taxa show the general features of segments 2 and 3 somewhat as in the Sciomyzoidea, but with some modifications. The distal articular surface of segment 2 is generally somewhat concave and encircled or almost so by the prominent rim, but the latter is often interrupted by a slight mid-dorsal notch, not by a long cleft as in most platystomatids and calyptrates. The conus is broad, almost uniformly sclerotized, rather short, and, with a little variation, more nearly bilaterally symmetrical than in most sciomyzoid taxa; it is tilted so as to retain its ventral prominence, often with a slight chin, but dorsally it is often scarcely raised above the general level of the articular surface. The distal foramen is vertically more or less elongate and tilted dorsally but not or only slightly laterally (as in Piophila ). The foraminal ring usually bears a dorsal and a ventral cusp (except in Herina ). The button occupies a dorsolateral position rather near the annular ridge. No taxon in this group is known to possess an encircling furrow (round the base of the conus) or caesti between the foraminal ring and annular ridge.
Segment 3 has usually a capacious basal hollow, but in Lonchaea it is reduced and divided by a ridge ( Fig. 132 View Figures 126–133 ). The basal stem is generally absent or only slightly indicated. The basal foramen is on a very slightly developed prominence near the margin of the basal hollow and usually faces ventrally, almost symmetrically.
The examples studied all have a three-segmented arista.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |