
Sibon noalamina, Lotzkat, Sebastian, Hertz, Andreas & Köhler, Gunther, 2012
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3485.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:DF69ABFD-AEAA-4890-899A-176A79C3ABA |
|
DOI |
https://doi.org/10.5281/zenodo.5617811 |
|
persistent identifier |
https://treatment.plazi.org/id/E1592540-366F-FF99-FF45-881AFCF2F8DF |
|
treatment provided by |
Plazi |
|
scientific name |
Sibon noalamina |
| status |
sp. nov. |
Sibon noalamina sp. nov.
Figs. 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3
Dipsas articulata: Köhler (2008: p. 219: Fig. 592) ; Stadler (2010); Carrizo (2010: in part, referring to Stadler 2010) Holotype. SMF 91539 (original field number SL 775; Figs. 1 View FIGURE 1 A–C, 2–3), adult male, from headwaters of Río Chiriquí Malí , approximately 6.4 km NW Fortuna dam (8.7891°N, 82.2155°W, 1050 m), Bosque Protector Palo Seco , Comarca Ngöbe-Buglé (formerly province of Bocas del Toro), Panama; collected by Andreas Hertz and Sebastian Lotzkat on 10 August 2010. GoogleMaps
Paratypes. SMF 90180 (original field number SL 494; Fig 1 View FIGURE 1 D), juvenile male, same locality as holotype; collected by Andreas Hertz and Sebastian Lotzkat on 29 October 2009 ; SMF 89550 (original field number LSt 015; Fig 1 View FIGURE 1 E), juvenile, from Cerro Mariposa near Alto de Piedra, approx. 3.5 km W of Santa Fé, 8.5001°N, 81.1170°W, 1260 m, province of Veraguas, Panama; collected by Sebastian Lotzkat and Andreas Hertz on 28 May 2008.
Diagnosis. Sibon noalamina differs from all described species of Sibon , and from all other Central American snail-eaters, in its slight keeling on the third to fifth dorsal row at midbody in adults, and, most obviously, in having only five supralabials, with the fifth and ultimate one being the only supralabial posterior to the orbit (versus two supralabials posterior to orbit in all other species of Sibon ) and exhibiting a peculiar shape: Its anterior portion is almost twice as high as the remaining supralabials, resembling the enlarged penultimate supralabial of other Sibon . Then it decreases in height towards the posterior portion that is about as high as the third supralabial, resembling the ultimate, usually moderately-sized, supralabial of other Sibon .
Additionally, its contrasting color pattern of complete dark rings on light background distinguishes S. noalamina from all Lower Central American Sibon except S. anthracops (Cope) , from which it differs by having 15 dorsal rows throughout the body (vs. 13 in S. anthracops ). At first sight, especially the more contrastingly colored juveniles of the new species might be confused with the coral snake-mimics Dipsas articulata , D. bicolor (Günther) , D. temporalis (Werner) , or D. viguieri (Bocourt) . From these, S. noalamina is distinguished by the presence of a mental groove (lacking in the genus Dipsas ), lower ventral counts (164–177 vs. 196–217 in D. articulata , 186–199 in D. bicolor , 170–208 in D. temporalis , and 190–203 in D. viguieri ), the unique supralabial condition (5 supralabials with only the ultimate, peculiarly shaped one posterior to orbit versus 9–10 supralabials in D. articulata , 10–11 in D. bicolor , 7–8 in D. temporalis , and 9–10 in D. viguieri , all shaped similarly, with three or more, rarely two, posterior to orbit), and fewer infralabials (6–7 infralabials versus 11–12 in D. articulata , 10–11 in D. bicolor , 8–13 in D. temporalis , and 9–11 in D. viguieri ). Furthermore, S. noalamina differs from D. temporalis in retaining the contrast between light and dark rings throughout body and tail ( Fig. 1 View FIGURE 1 ), whereas in D. temporalis the light portions grade into medium to dark brown posteriorly ( Fig. 5 View FIGURE 5 H).
Description of the holotype. Adult male ( Figs. 1 View FIGURE 1 A–C, 2–3), as indicated by everted hemipenes; SVL 385 mm, TL 161 mm (but tail incomplete); TOL 546 mm; HL 12.4 mm; HW 8.2 mm; SL 3.5 mm. Head distinct from neck; snout short, blunt in dorsal and lateral outline; rostral wider (2.1 mm) than high (1.3 mm), not extending posteriorly between internasals, its length visible from above one fifth length of median internasal suture (1.0 mm); internasals shorter (1.1 mm) than wide (1.7 mm), their length between one half and two-thirds length of prefrontal suture (1.9 mm); prefrontals shorter (2.5 mm) than wide (2.8 mm), their median suture (2.1 mm) about three-fifths length of frontal; prefrontals bordering eye above loreal; frontal longer (3.6 mm) than wide (3.0 mm), widest anteriorly, length almost two-thirds that of parietal, in contact with prefrontals, supraoculars, and parietals; supraoculars longer (3.4 mm) than wide (1.7 mm), nearly as long as frontal; parietals longer (5.8 mm) than wide (3.8 mm), widest anteriorly, their median suture (4.1 mm) slightly longer than frontal; parietals in contact with frontal, supraoculars, upper postoculars, anterior and posterior temporals, and nine nuchal scales; nasal divided, in contact with first two supralabials, loreal, prefrontal, internasal, and rostral; loreal single, longer (1.9 mm) than high (1.0 mm), its length more than one-half length of snout, and two-thirds length of eye (2.8 mm); preoculars absent; postoculars 2; temporals 1+2/2+2, all above fifth supralabial; supralabials five, with third and fourth bordering eye (enlarged anterior portion of ultimate supralabial separated from eye by lower postocular); fifth and ultimate supralabial conspicuously long (3.9 mm), longer than combined lengths of third and fourth supralabials, with a higher (1.7 mm) anterior and a lower (1.2 mm) posterior portion; pupil vertically elliptical; eye length one and a half times length of loreal; infralabials 6, first pair in contact behind mental; no postmental; infralabials 1–4 in contact with anterior chin shields; anterior chin shields paired, longer (2.8 mm) than wide (1.3 mm), longer than posterior chin shields; posterior chin shields longer (1.9 mm) than wide (1.4 mm), paired, in broad contact medially; mental groove present; ventrals 170; cloacal scute single; subcaudals 80 (but tail incomplete), divided; ventrals plus subcaudals 250 (but tail incomplete); dorsal scales mostly smooth, but slightly keeled on dorsal rows 3–5 around midbody, striate, in 15–15–15 rows, without apical pits or supracloacal ridges; vertebral row slightly enlarged.
The fully everted hemipenis ( Fig. 3 View FIGURE 3 ) is a stout unilobed capitate organ; the capitulum is covered by calyces bearing spinules; no naked pocket; the sulcus spermaticus is bordered by well-developed sulcal lips, bifurcates at the base of apex with the branches continuing onto the apex; truncus with two series of large recurved spines.
Coloration in life. Dorsal, lateral, and ventral surfaces of body and tail Pale Horn Color (92); body with 14, tail with eight broad Sepia (219) rings, body rings extending over 6–12 middorsal scales, narrowing towards and on venter; interspaces between these rings extending over 3–6 middorsal scales, speckled with Sepia (219) except for narrow portions immediately adjacent to the dark rings; tip of (incomplete) tail Sepia (119); dorsal and lateral surfaces of head Pale Horn Color (92), grading into Flesh Ocher (132D) anteriorly, heavily mottled with Sepia (219); parietals, frontal, and central portions of prefrontals almost completely Sepia (219), as well as the middorsal scales of the first two transverse rows of dorsals posterior to parietals, connecting Sepia (219) coloration to first body ring; ventral surface of head Pale Horn Color (92) mottled with Sepia (219), especially on infralabials; iris Sepia (119). Color after ten months in preservative (70% ethanol) is similar to that in life, apart from that all yellow and orange shades have faded.
Variation. The juvenile paratypes (SMF 89550, Fig. 1 View FIGURE 1 E; and SMF 90180, Fig. 1 View FIGURE 1 D) agree well with the holotype in terms of general morphology and pholidosis (even in their tails being incomplete), differing mostly in their lack of any keels on dorsal rows 3–5 and in their somewhat more contrasting coloration. Scale counts that differ from that of the holotype are as follows: ventrals 164 (SMF 89550), 177 (SMF 90180); subcaudals 93, 96 (both tails are incomplete); ventrals plus subcaudals 257, 273 (but both tails are incomplete); temporals 1+2/1+ 2 in both, anterior temporals of both sides entering orbit between upper and lower postoculars (SMF 89550); parietals bordered posteriorly by 8 nuchal scales in both; infralabials 6/7, 1– 5 in contact with first chin shield on left side (SMF 89550). Body measurements of the juvenile specimens are SVL 178 mm, TL 68 mm (SMF 89550), SVL 161 mm, TL 64 mm (SMF 90180).
As illustrated in Fig. 1 View FIGURE 1 , the coloration in life of the juveniles presents a stronger contrast than that of the adult holotype. Moreover, the paratypes show variation in the relative width of dark and light rings. The latter appear most narrow in the juvenile from Veraguas (SMF 89550), which was recorded as follows (modified from Stadler 2010): Body and tail with alternating, complete dark and light rings; broader rings Jet Black (89), light rings Chamois (123D), about half as wide as, and becoming lighter toward, the dark rings; dorsal and lateral ground color of head Jet Black (89); crescent-shaped lateral markings on posterior portion of head and small blotches on snout Chamois (123D); ventral surfaces of head and neck Chamois (123D) mottled with Jet Black (89); iris black. The color after 37 months in preservative (70% ethanol) is similar to that in life, apart from that all yellow and orange shades have faded.
Natural history notes. All three specimens were encountered at night on vegetation. The juvenile SMF 89550 ( Fig. 1 View FIGURE 1 E) from Cerro Mariposa, Veraguas, was moving through epiphytic liverworts growing on a small tree-trunk about 0.5 m above ground at 18:40 hrs, after a rainy afternoon. The habitat may be characterized as ridgetop cloud forest with abundant epiphytic vegetation. Annual precipitation is approx. 2500 mm, mean annual temperature approx. 20.2 °C, indicating the Premontane Wet Forest life zone according to the Holdridge (1967) classification. Other snail-eaters found at lower elevations on the slopes of Cerro Mariposa are Dipsas temporalis , Sibon annulatus , S. longifrenis , and S. nebulatus . A complete list of the herpetofauna documented around Cerro Mariposa was given by Stadler (2010), who also provided extensive data on climate and vegetation. Additionally, the herpetofauna of the general Santa Fé region was treated by Martínez and Rodriguez (1994 “1992”), Martínez et al. (1995 “ 1994 ”), and Carrizo (2010), and updated by Lotzkat et al. (2010).
Figure 1 View FIGURE 1 D shows the juvenile SMF 90180 as it was encountered at 19:40 hrs and 19.1 °C air temperature at the type locality: It was lying in loose coils upon a large leaf about 0.5 m above ground, in herbaceous secondary vegetation below the floodline of, and less than 10 m away from, a small stream. The holotype was found close to this stream while moving about in secondary forest, 2 m above ground at 23:00 hrs. The type locality is the valley of the mentioned stream, with rather undisturbed forest on the slope ascending northwards, minor clearings overgrown with grass and solitary trees directly by the riverside, and a more secondary forest on the slope ascending southward. Reptiles collected in this valley include Anolis capito Peters , A. humilis Peters , A. limifrons Cope , A. lionotus Cope , Lepidoblepharis xanthostigma (Noble) , Imantodes cenchoa (Linnaeus) , Sibon annulatus , and Bothrops asper ( Garman) . Annual total precipitation at the type locality is approx. 3000 mm, mean annual temperature approx. 20.6 °C, indicating the Premontane Wet Forest life zone according to the Holdridge (1967) classification. The datalogger recordings yielded a temperature range of 17.6–22.6 °C, with a mean of 19.4 °C and standard deviation ± 1.3 °C.
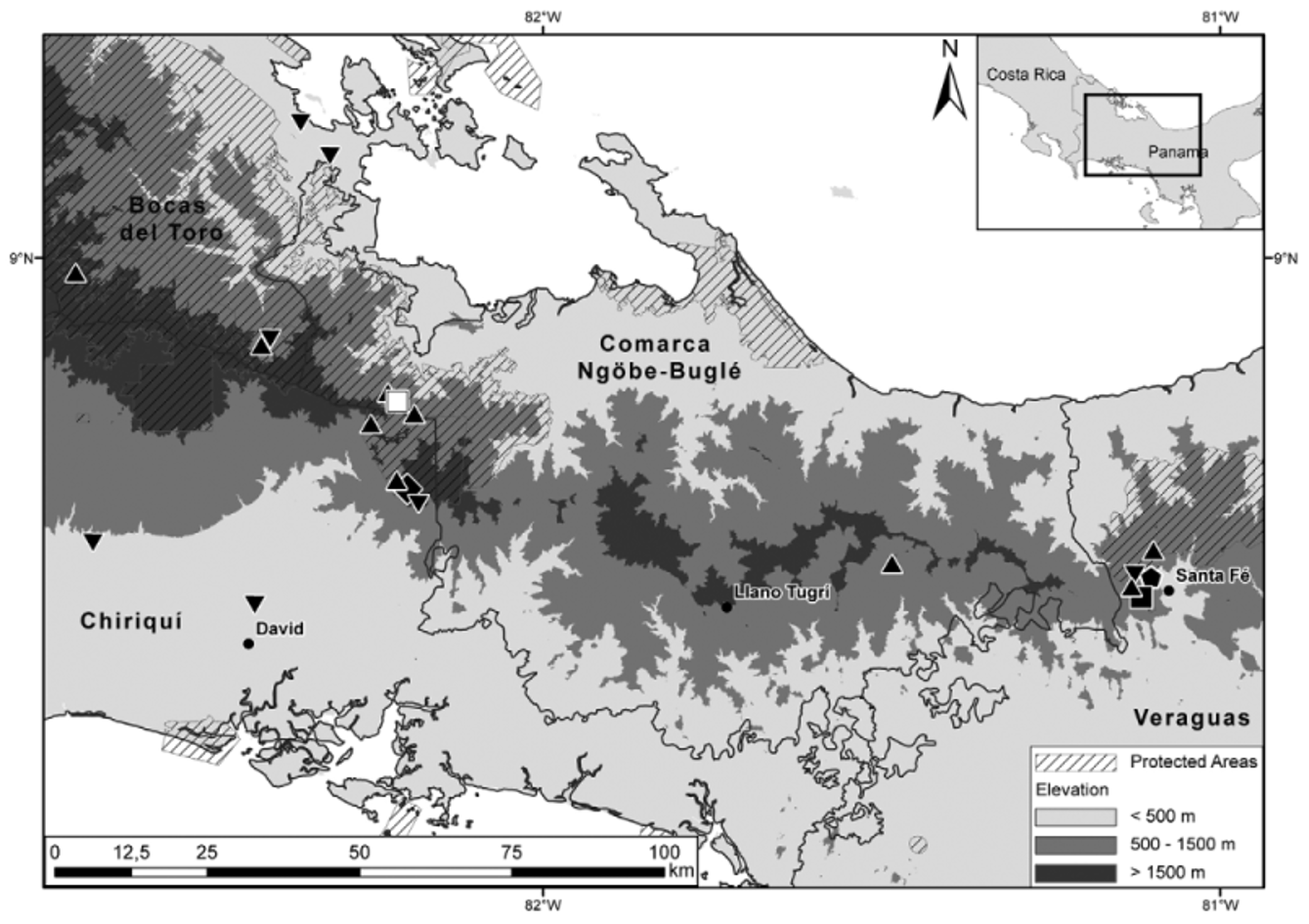
Geographic distribution. Sibon noalamina is hitherto only known from the highlands of western Panama ( Fig. 4 View FIGURE 4 ). The two known localities lie some 125 airline km apart, roughly at either end of the easternmost uninterrupted portion of the Cordillera Central, known as Serranía de Tabasará, which is approximately delimited to the west by the Fortuna depression and has its eastern limit in the region around Santa Fé. This indicates, as a minimum, a continuous distribution of the new taxon along the Caribbean versant of the Serranía de Tabasará, at premontane elevations between 1050 and 1260 m, most of it located in the Comarca Ngöbe-Buglé.
Etymology. The specific epithet is a contraction of the exclamation “no a la mina!”, Spanish for “no to the mine”, in the sense of “no mining”. This affirmation was and is used by members of the indigenous Ngöbe communities living in the Serranía de Tabasará in the course of their protests against mining interests aiming to exploit their territory, especially around Cerro Colorado. The specific name is given in recognition and support of the Ngöbe’s struggle to protect their territory and environment, which is home to the new species described herein and many others, from profit-driven destructive interventions.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.