
Pandinoides cavimanus ( Pocock, 1888 )
|
publication ID |
https://doi.org/ 10.1206/0003-0090-407.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.4630948 |
|
persistent identifier |
https://treatment.plazi.org/id/038C8783-8950-FF8E-BFB4-FB2BFB4FB89F |
|
treatment provided by |
Felipe |
|
scientific name |
Pandinoides cavimanus ( Pocock, 1888 ) |
| status |
|
Pandinoides cavimanus ( Pocock, 1888) View in CoL
Figures 1A, B View FIGURE 1 , 2 View FIGURE 2 , 3–9 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 ; table 1
Scorpio cavimanus Pocock, 1888: 247–249 (part); Pocock, 1896: 431, 432 (part), pl. XVIII, fig. 2, 2a.
Pandinus cavimanus: Kraepelin, 1899: 121 , 122 (part); 1908: 191, fig. 17; 1913: 183 (part); Birula 1915a: 51; Birula, 1915b: 26, 27 (part); Loveridge, 1925: 304, 305; Birula, 1927: 87, 88; Werner, 1934: 152, 278, fig. 116g; Werner, 1936: 185, 186 (part); Moriggi, 1941: 94 (part); Belfield, 1956: 43, 45; Probst, 1973: 327 (part); Kovařík, 1992: 186; 1998: 140 (part).
Pandinus cavimnus: Kraepelin, 1901: 270 .
Pandinus (Pandinoides) cavimanus: Vachon, 1974: 953 ; Lamoral and Reynders, 1975: 564 (part); El-Hennawy, 1992: 100, 136; Kovařík, 1997: 183; Fet, 2000: 468; Kovařík, 2002: 1, 18, 20 (part); 2003: 136, 148, 149, fig. 15, table 1 (part); 2009: 50–52 (part), 58, 59, 129 (part), table 3 (part), pl. K (fig. 2), figs. 403, 405.
Pandinoides cavimanus: Rossi, 2015a: 13 View in CoL (part), 50, figs. 8–10.
TYPE MATERIAL: Holotype ♂ ( BMNH 1863.66 ) [examined], TANZANIA: Tabora Prov.: Uyui Distr.: Central Africa , between the
center and Umyamuezi [04°59′S 033°10′E] and 5° to 7°S latitude, Captain Speke.
Fet (2000: 468) mistakenly listed two syntypes and Kovařík (2003: 149) erroneously designated one, BMNH 1863.66 from Umyamuezi, the lectotype (also see Kovařík, 2009: 58). Despite mentioning two specimens in the original description of Scorpio cavimanus, Pocock (1888: 249) explicitly designated one as the type: “I have seen two specimens … one (dried) brought from Kilima- Njaro by Mr. M. J. Jackson; the other, which, being preserved in spirit of wine, I have selected as the type, brought by Capt. Speke from Umyamuezi.”
DIAGNOSIS: Pandinoides cavimanus may be separated from P. duffmackayi , sp. nov., and P.militaris , as follows. Pandinoides cavimanus is usually brownish black (fig. 1A, B), rarely reddish brown, in color whereas P. duffmackayi , sp. nov., is reddish brown to reddish black (fig. 1C) and P. militaris , yellowish brown (fig. 1D) to reddish brown. The interocular and circumocular surfaces of the carapace are sparsely and finely granular anterior to the median ocular tubercle, along the median longitudinal sulcus, and often on the frontal lobes in P. cavimanus (fig. 4A, C), but smooth, or nearly so, in P. duffmackayi , sp. nov. (fig. 11A, C), and smooth or with a few granules anterior to the ocular tubercle in P. militaris (fig. 18A, C). The metasoma of P. cavimanus is 52%–56% (♂) and 49%–53% (♀) of total length, the summed lengths of segments IV and V, 115%–130% (♂), 107%– 120% (♀) of carapace length (table 1), whereas the metasoma of P. duffmackayi , sp. nov., is 49%–53% (♂) and 46%–49% (♀) of total length, the summed lengths of segments IV and V, 105%–116% (♂) and 91%–110% (♀) of carapace length (tables 2, 3), and the metasoma of P. militaris is 46%–52% (♂) and 42%–50% (♀) of total body length, the summed lengths of segments IV and V, 98%–117% (♂) and 87%–105% (♀) of carapace length (tables 4, 5). The dorsal surfaces of metasomal segments I–IV in the male are finely and sparsely granular in P. cavimanus (fig. 9A) but smooth in P. duffmackayi , sp. nov. (fig. 16A), and P. militaris (fig. 23A). The ventral surface of the telson vesicle of the male bears obsolete carinae, each comprising isolated spiniform granules, restricted to the anterior half or third, in P. cavimanus (fig. 9B, C), whereas the vesicle is smooth or nearly so in P. duffmackayi , sp. nov. (fig. 16B, C), and bears distinct carinae, each comprising prominent spiniform granules, extending its entire length, in P. militaris (fig. 23B, C). The proximal lobe on the pedipalp chela fixed finger of the adult male is slightly smaller than the medial lobe, creating a moderate gap proximally between the fixed and movable fingers, when closed in P. cavimanus (fig. 6A), whereas the proximal lobe is similar in size to the medial lobe, creating little to no gap proximally between the fingers, when closed in P. duffmackayi , sp. nov. (fig. 13A), and it is vestigial, much smaller than the medial lobe, or absent, creating a prominent gap proximally between the fingers, when closed in P.militaris (fig. 20A).
Based on unpublished genetic data, P. cavimanus is most closely related to P. duffmackayi , sp. nov., from which it may be further separated as follows. Pandinoides cavimanus is larger, with total adult body length 87–111 mm, carapace length 15–18 mm and pedipalp chela length 21–29 mm (table 1), than P. duffmackayi , sp. nov., with total adult body length 60–78 mm, carapace length 10–13 mm and chela length 14–19 mm (tables 2, 3). Pandinoides cavimanus bears 83–91 trichobothria on the pedipalp, with 48–55 trichobothria on the patella, including 31–37 in the v series, whereas P. duffmackayi , sp. nov., bears 74–82 trichobothria on the pedipalp, with 39–45 trichobothria on the patella, including 22–28 in the v series. The pectinal tooth count of P. cavimanus , with 14 or 15, usually 14 (♂) and 13 or 14, usually 13 (♀), is higher than that of P. duffmackayi , sp. nov., with 11–13, usually 12 (♂) and 10–12, usually 11 (♀). The prolateral t and st macrosetae of basitarsi III and IV are spiniform in P. cavimanus (fig. 8C, D), whereas t and st are setiform on III, and t setiform and st spiniform on IV in P. duffmackayi , sp. nov. (fig. 15C, D).
Pandinoides cavimanus has often been confused with P.militaris due to their similar size and high trichobothrial counts, but may be separated from the latter as follows. The carapace is less compressed, dorsoventrally, in P. cavimanus (fig. 4A, C) than in P. militaris (fig. 18A, C). The median ocular tubercle is posteromedial, its distance from the anterior carapace margin 51%– 56% of the carapace length in P. cavimanus (table 1), but medial, its distance from the anterior carapace margin 46%–52% of the carapace length, in P. militaris (tables 4, 5). The ventrosubmedian carinae on metasomal segment IV are distinct, costate in P. cavimanus (fig. 9C), but obsolete, granular in P.militaris (fig. 23C). The chela manus of P. cavimanus is broader, the length along the retroventral carina 56%–63% (♂) and 58%–67% (♀) of the width, and more convex, the height 59%–73% (♂) and 66%–76% (♀) of the width, than the manus of P.militaris , in which the length along the retroventral carina is 64%–76% (♂) and 62%–72% (♀) of the width, and the height 49%– 68% (♂) and 64%–73% (♀) of the width. The chela fixed finger of P. cavimanus arises abruptly from the manus and is wider proximally (figs. 6A, 7) than that of P.militaris , which arises more gradually from the manus (figs. 20A, 21). The chela fingers and distal surfaces of the manus are moderately (♂) to sparsely (♀) setose in P. cavimanus (fig. 1B), but densely (♂) to moderately (♀) setose in P. militaris (fig. 1D). Pandinoides cavimanus bears 31–35 trichobothria on the pedipalp chela, including 7–10 in the V series, whereas P.militaris bears 33–36 trichobothria on the chela, including 9–12 in the V series. The angle of the first proximal median lamella (scape) of the pecten is smaller (more acute), especially in the female, of P. cavimanus (fig. 4B, D), compared with P. militaris (fig. 18B, D).
REDESCRIPTION: The following redescription supplements the original description by Pocock (1888) and is based on the material examined.
Total Length: Adult medium, maximum length, measured from anterior margin of carapace to tip of aculeus, 106 mm (104–111 mm, n = 4) (♂), 95 mm (87–105 mm, n = 7) (♀) (table 1).
Color: Chelicerae, dorsal surfaces bicolored, proximal three-quarters of manus dorsal surface sparsely infuscate, paler than carapace and densely infuscate distal quarter of manus dorsal surface and fingers. Carapace, tergites, metasoma, telson, pedipalp trochanter, femur, patella pro- and retrolateral surfaces, chela fingers, and legs dorsal and retrolateral surfaces entirely infuscate, uniformly dark brownish black; pedipalp patella ventral surface and chela manus dorsal, lateral, and ventral surfaces entirely infuscate but slightly paler, dark maroon. Coxosternal region, sternites, and legs, prolateral and ventral surfaces predominantly infuscate, paler, dark olive-brown with maxillary lobes darker, and leg tibia, basitarsus, and telotarsus paler than femur and patella. Genital opercula and pectines, immaculate, uniformly pale cream.
Chelicerae: Movable finger, prodistal (ventral) and retrodistal (dorsal) teeth unequal, retrodistal tooth considerably smaller than prodistal tooth, aligned longitudinally and not opposable. Fingers and manus, proventral surfaces, with long, dense vestiture of macrosetae. Coxae, prodorsal surfaces without stridulatory setae (scaphotrix); promedian surfaces without chemoreceptive lamelliform setae (trichocopae).
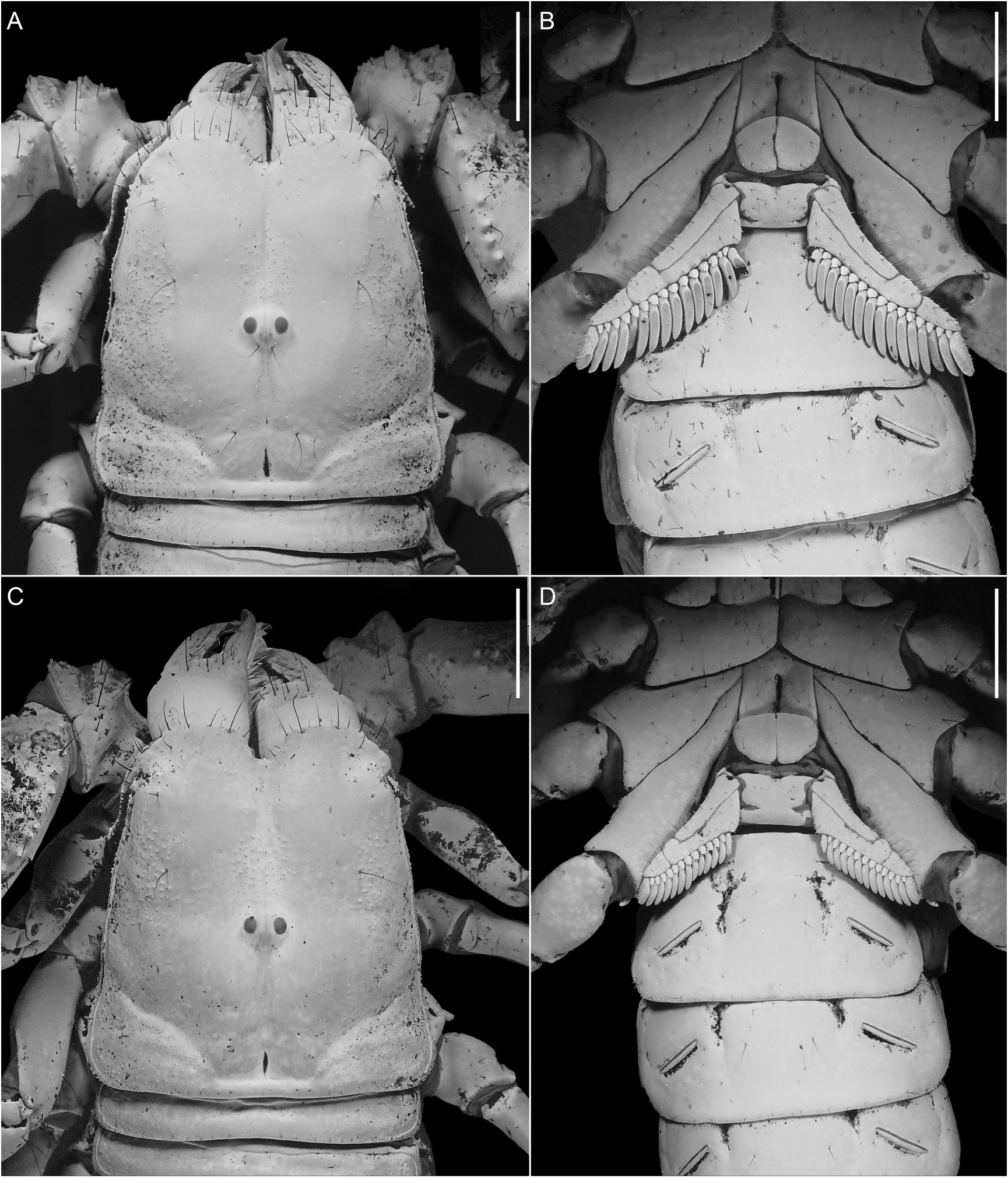
Carapace: Anterior width of posterior width, 72% (69%–73%, n = 4) (♂), 71% (66%–76%, n = 7) (♀); posterior width of length, 99% (97%– 101%, n = 4) (♂), 98% (91%–104%, n = 7) (♀) (table 1). Three pairs of lateral ocelli, anterior pairs larger, facing anteriorly, separated by distance equal to width of ocellus from smaller posterior pair, facing posteriorly. Median ocelli slightly larger than anterior pairs of lateral ocelli, distance between ocelli equal to or greater than width of ocellus; median ocular tubercle situated posteromedially, distance from anterior carapace margin 54% (52%–56%, n = 4) (♂), 54% (51%–56%, n = 7) (♀) of carapace length (table 1). Anterior carapace margin with deep median notch (fig. 4A, C); with small median projection (epistome); without median depression; rostrolateral margin without distinct notch next to posterior lateral ocelli. Median longitudinal sulcus narrow, suturiform; continuous from median notch to interocular sulcus; with obsolete anterior furcation; without anterocular depression. Median ocular tubercle raised, superciliary carinae higher than ocelli, not extended anteriorly or posteriorly. Interocular sulcus present. Circumocular depressions completely encircling median ocular tubercle, converging anteriorly. Posteromedian and posteromarginal sulci deep. Paired mediolateral and posterolateral sulci shallow. Circumocular depressions completely encircling median ocular tubercle, converging anteriorly. Posteromedian and posteromarginal sulci deep. Paired mediolateral and posterolateral sulci shallow. Median longitudinal suture continuous from median notch to median ocular tubercle, equally strong along entire length; not extending to anterior margin of carapace, terminating at or posterior to median notch. Anterior furcated sutures obsolete. Anterocular furcated sutures absent. Interocular suture present, slender. Posterior sutures present but less distinct than median longitudinal and interocular sutures, converging on median ocular tubercle; connected anteriorly to posterior furcations of interocular suture and extending anteriorly beyond median ocular tubercle. Interocular and circumocular surfaces sparsely and finely granular anterior to median ocular tubercle, along median longitudinal sulcus, and often on frontal lobes (fig. 4A, B). Anterolateral, mediolateral and posterolateral surfaces uniformly finely granular, more coarsely and densely so in male. Posteromedian surfaces smooth.
Pedipalps: Femur width of length, 53% (52%– 53%, n = 4) (♂), 57% (48%–62%, n = 7) (♀) (table 1). Retrodorsal carina distinct, comprising several spinform granules; more strongly developed than prodorsal carina. Dorsomedian carina vestigial, reduced to prominent granule demarcated by conspicuous macroseta. Dorsal secondary carina absent. Prodorsal carina obsolete, comprising isolated subspiniform granules. Promedian carina distinct, comprising row of spiniform granules (several demarcated by conspicuous macrosetae), oriented diagonally between prodorsal and proventral carinae. Proventral carina distinct, costate-granular, more strongly developed than retroventral carina. Ventromedian, secondary accessory, and retroventral carinae absent. Retromedian carinae absent, indicated only by macrosetal rows. Prodorsal intercarinal surfaces with scattered granules; other intercarinal surfaces smooth. Patella width of length, 45% (42%–47%, n = 4) (♂), 44% (42%–47%, n = 7) (♀) (table 1). Dorsal surface convex, dorsomedian carina dorsal to plane of retrodorsal carina, obsolete, costate (fig. 5A). Retrodorsal and retromedian carinae absent. Retroventral carina obsolete, costate (fig. 5B). Promedian carina obsolete, costate, demarcated by conspicuous macroseta; anterior process absent. Other carinae absent. Intercarinal surfaces smooth. Chela short, broad, base of fixed finger arising abruptly from manus (figs. 6A, 7); manus, height of width, 66% (59%–73%, n = 4) (♂), 61% (56%–63%, n = 7) (♀); length along ventroexternal carina of width, 61% (56%–63%, n = 4) (♂), 62% (58%–67%, n = 7) (♀); length along ventroexternal carina of length movable finger, 52% (50%–57%, n = 4) (♂), 57% (53%–60%, n = 7) (♀) (table 1). Dorsomedian carina obsolete, except proximal to base of fixed finger, distinct, costate. Dorsal secondary, subdigital, retromedian, retrolateral secondary and secondary accessory carinae absent. Digital carina absent, except proximal to base of fixed finger, obsolete costate. Retroventral and ventromedian carinae distinct, costate; retroventral carina more pronounced and projecting ventral to plane of ventromedian carina (fig. 6B). Proventral and promedian carinae obsolete, granular, each indicated by prominent macroseta. Prodorsal carina absent, but indicated by prominent macroseta. Chela moderately (♂) to sparsely (♀) setose on fingers and distally on manus. Manus, retrodorsal surface with (adult ♂) or without (♀, immature stages) marked concave depression at base of fixed finger (figs. 6A, 7), predominantly smooth proximally with shallow, anastomosing granules distally; retrolateral surfaces shallowly granular; ventral intercarinal surface smooth; prolateral intercarinal surfaces predominantly smooth, with scattered spiniform granules dorsally and distally. Fixed and movable fingers, prolateral intercarinal surfaces finely granular, retrolateral intercarinal surfaces predominantly smooth (♂) or finely granular (♀); median denticle rows each with six enlarged retrolateral denticles (including terminal denticle), proximal three retrolateral denticles on fixed finger, and proximal three (♀) or second and third most proximal (♂) retrolateral denticles on movable finger situated on lobes; fixed finger with proximal lobe, similar in size (♀, immatures) or slightly smaller than medial lobe (adult ♂); movable finger (adult ♂) without proximal lobe, with median lobe markedly more pronounced than other lobes, and with correspondingly well-developed notch in fixed finger; moderate gap proximally between fingers, when closed (adult ♂); terminal denticles of fingers interlocking unevenly when closed, movable finger displaced to exterior; distinct notch near tip of fixed finger to accommodate terminal denticle of movable finger.
Trichobothria: Neobothriotaxic major, Type C, with the following segment totals (n = 25; table 1): femur, 3 (1 d, 1 i, 1 e); patella, 52 (48– 55): 2 d, 1 i, 34 (31–37) v, 14 (14 or 15) e, usually comprising 3 et, 2 est, 2 em, 2 esb, 5 eb; chela, 33 (31–35), manus, 21 (19–22), comprising 2 D, 10 E, 9 (7–10) V; fixed finger, 12 (11–13), comprising 4 d, 4 e, 4 (3–5) i (figs. 5–7). Total count of trichobothria per pedipalp: 87 (83–91). Femur, i situated on dorsal surface. Patella, d 2 situated on dorsomedian carina, slightly closer to d 1 than to i. Chela, distance et–est ca. half distance est–esb; distance est–esb greater than half distance esb–eb; est aligned with dst.
Legs: First pair of maxillary lobes (coxapophyses) tapering anteriorly, longer than and encircling second pair. Stridulatory organs, comprising “rasp” (granular tubercles) and “scraper” (stridulatory setae or scaphotrix), present on opposing surfaces of coxae of pedipalps and first pair of legs, respectively. Legs acarinate. Femora, patellae, and tibiae, pro- and retrolateral surfaces each with scattered setiform macrosetae. Tibiae, prolateral surfaces, without spiniform macrosetae; I and II, retrolateral surfaces, usually each with two spiniform (t, st) macrosetae; III and IV, retrolateral surfaces, without spiniform macrosetae. Basitarsi I–IV, prolateral pedal spurs present (fig. 8); retrolateral pedal spurs absent; retrolateral margins similar, unmodified, rounded; I and II, dorsoventrally compressed, retrolateral margin produced into bladelike edge, III and IV terete, retrolateral margin unmodified, rounded. Basitarsi, pro- and retrolateral surfaces, each with microsetae, scattered long and short setiform macrosetae, and spiniform macrosetae, more numerous on I and II than III and IV. Basitarsi, spiniform macrosetae, I, retrolateral: t, st (polymorphic), sb; retroventral: t; proventral: t, st; II, retrolateral: t, st (polymorphic), sb; retroventral: t; proventral: t, st; III, retrolateral: t, sb; retroventral: t; proventral: t, st; prolateral: t, st; IV, retrolateral: t; retroventral: t; proventral: t, st; prolateral: t, st. Telotarsi I–IV short, stout, and distally broadened in dorsal and lateral views. Laterodistal lobes rounded. Dorsomedian lobes approximately equal to laterodistal lobes; each terminating in single setiform macroseta. Telotarsi, pro- and retrolateral surfaces, each with scattered microsetae and setiform macrosetae; I and II, pro- and retrolateral surfaces, in addition with two or more, dense brushlike rows of long to very long setiform macrosetae. Telotarsi each with pro- and retroventral rows of spiniform macrosetae, two of which are inserted on laterodistal lobes; macrosetal counts in pro- and retroventral rows equal on telotarsi I–IV, 3 and 4 (4 or 5) (n = 25), respec- tively (table 1). Telotarsal ungues short, curved; equal on telotarsi I and II, subequal on III and IV.
Sternum: Shape subpentagonal (fig. 4B, D). Median longitudinal sulcus shallow anteriorly, deep and narrow posteriorly.
Genital Operculum: Genital opercula suboval, completely divided longitudinally, partially overlapping, genital papillae present (♂, fig. 4B); subcordate, fused, genital papillae absent (♀, fig. 4D).
Hemispermatophore: Lamelliform, with complex, folded capsule and accessory distal lobe protruding between articular suture and distal lobe (hook). Distal lamina with distal crest truncate, unfolded.
Pectines: Distal edge reaching past distal edge of coxa IV but not reaching to distal edge of trochanter IV (♂, fig. 4B) or almost reaching distal edge of trochanter IV (♀, fig. 4D). First proximal median lamella (scape) of each pecten with mesial margin obtusely angular, greater than 90° but less than 180°, and devoid of teeth in proximal 19% (14%–23%, n = 4) (♂) or 24% (17%– 31%, n = 7) (♀) of prolateral margin (table 1). Pectinal teeth straight and elongate (♂) or shorter and curved (♀); tooth count, 14/14 (13– 15/14–15, n = 4) (♂), 13/13 (12–14/12–14, n = 7) (♀). Fulcra smooth proximally but densely setose (microsetae only) distally.
Mesosoma: Tergites each with shallow pair of submedian depressions and obsolete median carina. Pretergites smooth and glabrous. Posttergites smooth and glabrous anteromedially, densely and coarsely granular posteromedially and laterally (♂) or smooth and glabrous medially and anterolaterally, sparsely and coarsely granular posterolaterally (♀). Sternites IV–VI, each with paired longitudinal depressions prolateral to spiracles, absent on VII. Surface, sternites III–VII, smooth; VII, with paired, costate ventrosubmedian and ventrolateral carinae, without posteromarginal carina. Sternite VII, length of width, 52% (44%–56%, n = 4) (♂), 56% (47%– 69%, n = 7) (♀) (table 1).
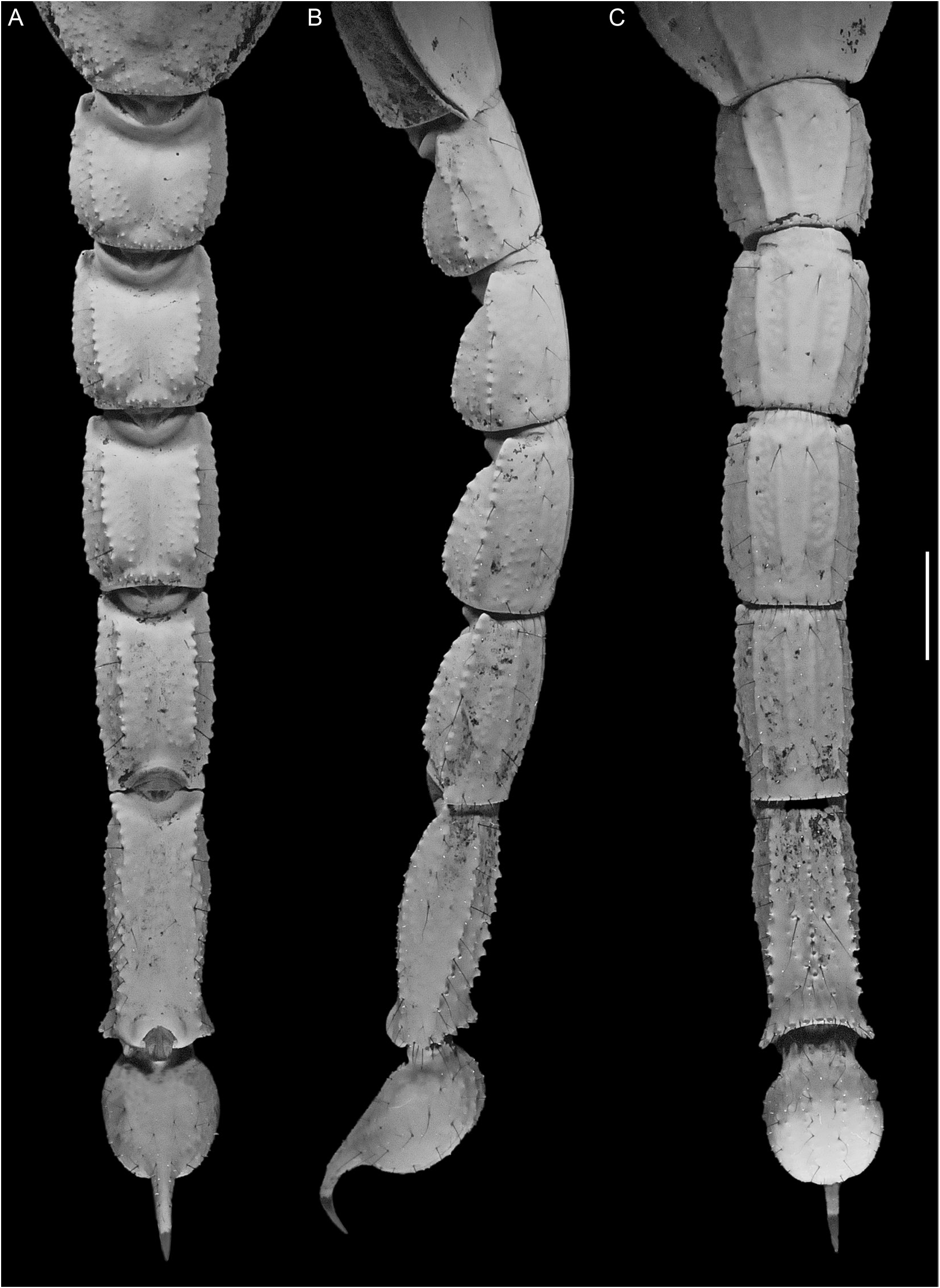
Metasoma and Telson: Metasomal segments I–V progressively increasing in length, decreasing in width; segment V, width of segment I width, 66% (64%–68%, n = 4) (♂), 66% (64%– 68%, n = 7) (♀) (table 1). Metasoma robust; width of length, segment I, 93% (90%–95%, n = 4) (♂), 91% (89%–93%, n = 7) (♀); II, 78% (74%–81%, n = 4) (♂), 80% (76%–86%, n = 7) (♀); III, 68% (65%–73%, n = 4) (♂), 69% (60%–77%, n = 7) (♀); IV, 55% (52%–57%, n = 4) (♂), 58% (52%–65%, n = 7) (♀); V, 43% (39%–48%, n = 4) (♂), 43% (40%–45%, n = 7) (♀). Telson vesicle, width of metasomal segment V, width, 111% (105%–115%, n = 4) (♂), 97% (87%–107%, n = 7) (♀); enlarged (♂), globose, height of length, 62% (58%–66%, n = 4) (♂), 62% (57%–65%, n = 7) (♀); dorsal surface flat, ventral surface evenly curved. Aculeus relatively short, strongly curved, length of vesicle length, 32% (31%–34%, n = 4) (♂), 37% (33%– 40%, n = 7) (♀). Length, metasoma and telson, of total length, 54% (52%–56%, n = 4) (♂), 51% (49%–53%, n = 7) (♀). Dorsosubmedian carinae, segments I–IV, distinct, complete, costategranular, posterior spiniform granules obsolete; V, absent (fig. 9A). Dorsolateral carinae, segments I–V, distinct, complete, costate-granular on I–IV, granular on V. Median lateral carinae, segment I, obsolete, granular, reduced to posterior two-thirds, II–V, absent, demarcated by macroseta at posterior margin on II–IV and near anterior margin on V (fig. 9B). Ventrolateral and ventrosubmedian carinae more strongly developed on segments I and II than on segments III and IV (fig. 9C). Ventrolateral carinae, segments I–IV, distinct, complete, costate on I–III, costate-granular on IV; V, distinct, complete, diverging posteriorly, comprising subspiniform granules, terminal granule similar in size to preceding granules. Ventrosubmedian carinae, segments I–IV, distinct, complete, costate; V, vestigial, each reduced to discontinuous row of spiniform granules, demarcated by macrosetae. Ventromedian carina, segment V, comprising single row of spiniform granules, unmodified posteriorly. Ventral surface, lateral aspect, segment IV, shallowly convex. Anal arch, segment V, dorsal carina, costate; ventral carina comprising subspiniform granules. Dor- sal and lateral intercarinal surfaces, segments I–IV, finely and sparsely granular; V, smooth. Ventral intercarinal surfaces, segments I–IV, smooth; V with scattered granules. Telson vesicle, dorsal and lateral surfaces smooth; ventral surface with four obsolete longitudinal carinae, each comprising isolated spiniform granules, restricted to anterior half or third of vesicle.
Geographical Variation: All except three specimens examined were dark brownish black in color. One specimen with an indefinite locality in “East Africa” (BMNH) and two of the five male specimens labelled “Ugogo district, German East Africa ” (BMNH 1914.7.3.3– 7) were reddish brown in color, resembling P. militaris .
Ontogenetic Variation: Immature males and females closely resemble each other, and adult females, in general morphological features, but can be sexed by examination of the pectines and genital operculum. Males acquire secondary sexual characters in the final instar. In the specimens examined for this study, sexual maturity was determined by the presence of a marked concave depression in the retrodorsal surface of the pedipalp chela manus, at the base of the fixed finger in males (fig. 6A) and a fully developed ovariuterus or the gravid condition in females.
Sexual Dimorphism: The characters of primary external sexual dimorphism are the undivided genital operculum, which opens in a single flap in the female (fig. 4D), compared with the two unconnected sclerites, which open independently and cover a pair of genital papillae in the male (fig. 4B). Additionally, males possess larger pectines (fig. 4B, D) with a greater number of pectinal teeth (table 1). The most obvious secondary sexual characters observed in adult males, compared with adult females and juveniles of both sexes, concern the modifications of the pedipalp chela: a marked concave depression is present in the retrodorsal surface of the manus, at the base of the fixed finger; the proximal lobe of the movable finger is absent; the median lobe of the movable finger is markedly more pronounced than the other lobes; and a moderate gap is present proximally between the fingers, when closed (fig. 6A).
DISTRIBUTION: Pandinoides cavimanus appears to be endemic to Tanzania (fig. 2). Records from Kenya are referable to P. duffmackayi , sp. nov., or P. militaris , those from Ethiopia or Somalia, including the (immature) female on the cover of Kovařík (2009), to P. militaris . Records of P. cavimanus from northeastern Tanzania, including Kilimanjaro ( Pocock, 1888), Arusha ( Werner, 1936), and Naberera ( Kovařík, 2009) are referable to P. duffmackayi , sp. nov. Most of the known locality records of P. cavimanus occur in the central part of Tanzania, in the Dodoma, Iringa, Shinyanga, and Singida provinces. Records from the eastern shore of Lake Tanganyika and Bukoba on the western shore of Lake Victoria/Nyanza ( Kraepelin, 1913; Birula, 1915b) await confirmation. This species does not occur in the northeast of the Democratic Republic of Congo (formerly Zaïre), as stated by Kovařík (1998, 2009) and Fet (2000), nor in Mozambique, as stated by Kovařík (1998).
ECOLOGY: The known locality records of P. cavimanus occur in arid savannah, dominated by Acacia Mill. and Commiphora Jacq. at 1010–1120 m elevation. Most specimens, for which data are available, were excavated from burrows during the day. Pandinoides cavimanus is fossorial, constructing burrows in compacted clayey to sandyloam soils. It is allopatric with P. militaris and parapatric with P. duffmackayi , sp. nov., and Pandinurus viatoris ( Pocock, 1890) , but sympatric with another scorpionid, Opistophthalmus boehmi (Kraepelin, 1896) .
CONSERVATION: This species is protected in the Ruaha National Park.
REMARKS: Pocock (1888: 249) noted that: “The specimen from Kilima-Njaro is smaller and slightly less granular than the [type] from Umyamuezi.” The smaller specimen is conspecific with P. duffmackayi .
In discussing the differences in coloration between P. exitialis and P. gregoryi , which Kovařík (2003) mistakenly synonymized with the former, Kovařík (2009: 55) remarked that:
Most of Pandinus species are based on unqiue specimens. While I studying [sic] the holotype of P. gregoryi , I was convinced that P. gregoryi is a synonym of P. exitialis (see Kovařík, 2003: 151). Subsequently I had an opportunity to study and breed both populations, P. exitialis from southern Somalia and P. gregoryi from central Kenya [ P. exitialis has since also been recorded from central Kenya (L. Prendini, unpublished data)], and can state that they are similar but nevertheless separate species which can be distinguished by granulation on dorsal side of the pedipalp manus – in P. gregoryi tuberculate but without conical granules of equal size, which in contrast characterize P. exitialis . Coloration is also different, especially the pedipalp chela is much lighter in P. exitialis than in P. gregoryi . However, differences in color should not be regarded as a decisive taxonomic criterion, which can be well demonstrated e.g. on the example of P. cavimanus , whose coloration is very variable in both juveniles and adults throughout the geographic range of this species.”
Whereas it is indeed true that color may vary both within and among conspecific populations of some scorpion species, e.g. Hadrurus arizonensis Ewing, 1928 , each case must be assessed on its own merits. Variation in color pattern (rather than color itself) is often both consistent and congruent with variation in other morphological (and indeed, genetic) characters, as described by Kovařík (2009) in the closely related P. exitialis and P. gregoryi , and the three closely related species of Pandinoides covered in the present contribution. Valid taxa, e.g., P. gregoryi and P.militaris , may be mistakenly synonymized when differences in coloration are dismissed out of hand without detailed investigation of other characters.
ADDITIONAL MATERIAL EXAMINED: 1 ♂ ( NM 18769 ). East Africa, 1 ♂ ( BMNH). TANZANIA: Imported for pet trade: 2 ♂ ( AMNH), 1 ♀ ( AMCC [ LP 3250 ]), 1 ♀ ( AMNH), [leg] ( AMCC [ LP 3259 ]), ix.2002, ex R. MacInnes , 1 subad. ♀ ( AMNH), [leg] ( AMCC [ LP 2161 ]), vii.2007, ex J. Huff, 1 ♂ ( AMCC [ LP 7084 ]), iii.2008, ex A. Tietz, 1 subad. ♀ ( AMNH), [pedipalp] ( AMCC [ LP 8271 ]). Ugogo district , German East Africa , Dr. E.J. Baxter, 5 ♂ ( BMNH 1914.7.3.3-7), 1 ♀, 1 subad. ♂ ( BMNH 1914.7.3.8–9). Dodoma Prov.: Dodoma Distr.: Dodoma [06°10′S 035°45′E], G.E.A., 8.xii.1918, A. Loveridge, 2 ♂ ( MCZ 15519), 1 subad. ♀, 4 juv. ( MCZ 15513), 1 ♂ ( BMNH 1923.11.8.9) GoogleMaps ; Dodoma, brought in by dealer, 1 ♂, 1 juv. ♂ ( ZMUC) ; Mapinduzi Village, Dodoma region [06°11′S 035°45′E], ix.2006, J. Beraducci, found in ground, 3 ♂, 3 ♀, 2 subad. ♂, 1 subad. ♀ ( AMNH), 1 juv. ♀ ( AMCC [ LP 8918 ]), x.2006, J. Beraducci, 1 ♂, 3 ♀, 1 subad. ♂ ( AMNH). Kongwa Distr.: Kongwa [06°12′S 036°25′E], 21.iv.1918, A. Loveridge, 1 ♀ ( MCZ 15522). Mpwapwa Distr. GoogleMaps : Mpapua [Mpwapwa, 06°21′S 036°29′E], 1899, 1 ♂ ( SAM 5111 View Materials ex ZMH). Iringa Prov.: Iringa Distr.: Ruaka [Ruaha] National Park [07°40′S 034°52′E], 1979, K.M. Howell, 1 ♂ ( MRAC 159.373 View Materials ). Singida Prov.: Manyoni Distr. GoogleMaps : Ikikuyu [Kikuyu, 05°52′S 035°04′E], 12.ii.1923, A. Loveridge, 1 ♂, 1 subad. ♂ ( MCZ 15514), 1 ♂, 1 juv. ♂ ( MCZ 15518), 1.xi.1923, A. Loveridge, 1 ♂ ( MCZ 15521), 12.xi.1923, A. Loveridge, 1 juv. ♂ ( MCZ 15516) ; Kikuyu , Dodoma, 21.xii.1929, A. Loveridge, 2 ♂, 1 ♀ ( MCZ 15515) .
ERRONEOUS RECORDS: Florida, U.S.A., New Bold’s 5-10, Clubmont (introduction), vii.1965, University of Florida Collections, 1 ♂ (FSCA). Stanley Falls [Tshopo Prov., Democratic Republic of Congo], 00°30′N 025°12′E, 1 ♂ (MRAC 4669). Based on the known localities, the record from Stanley Falls is assumed to be a labeling error.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pandinoides cavimanus ( Pocock, 1888 )
| Prendini, Lorenzo 2016 |
Pandinoides cavimanus: Rossi, 2015a: 13
| Rossi, A. 2015: 13 |
Pandinus (Pandinoides) cavimanus:
| Kovarik, F. 2002: 1 |
| Fet, V. 2000: 468 |
| Kovarik, F. 1997: 183 |
| El-Hennawy, H. K. 1992: 100 |
| Lamoral, B. H. & S. C. Reynders 1975: 564 |
Pandinus cavimnus:
| Kraepelin, K. 1901: 270 |
Pandinus cavimanus: Kraepelin, 1899: 121
| Kovarik, F. 1998: 140 |
| Kovarik, F. 1992: 186 |
| Probst, P. J. 1973: 327 |
| Belfield, W. 1956: 43 |
| Moriggi, M. 1941: 94 |
| Werner, F. 1936: 185 |
| Werner, F. 1934: 152 |
| Birula, A. A. 1927: 87 |
| Loveridge, A. 1925: 304 |
| Birula, A. A. 1915: 51 |
| Birula, A. A. 1915: 26 |
| Kraepelin, K. 1899: 121 |
Scorpio cavimanus Pocock, 1888: 247–249
| Pocock, R. I. 1896: 431 |
| Pocock, R. I. 1888: 249 |