
Scarabaeus (Pachysoma) hippocrates ( MacLeay, 1821 )
|
publication ID |
https://doi.org/ 10.1080/713834683 |
|
persistent identifier |
https://treatment.plazi.org/id/03A60308-FFEA-320D-FE1C-FBABFD3987B3 |
|
treatment provided by |
Carolina |
|
scientific name |
Scarabaeus (Pachysoma) hippocrates ( MacLeay, 1821 ) |
| status |
|
Scarabaeus (Pachysoma) hippocrates ( MacLeay, 1821)
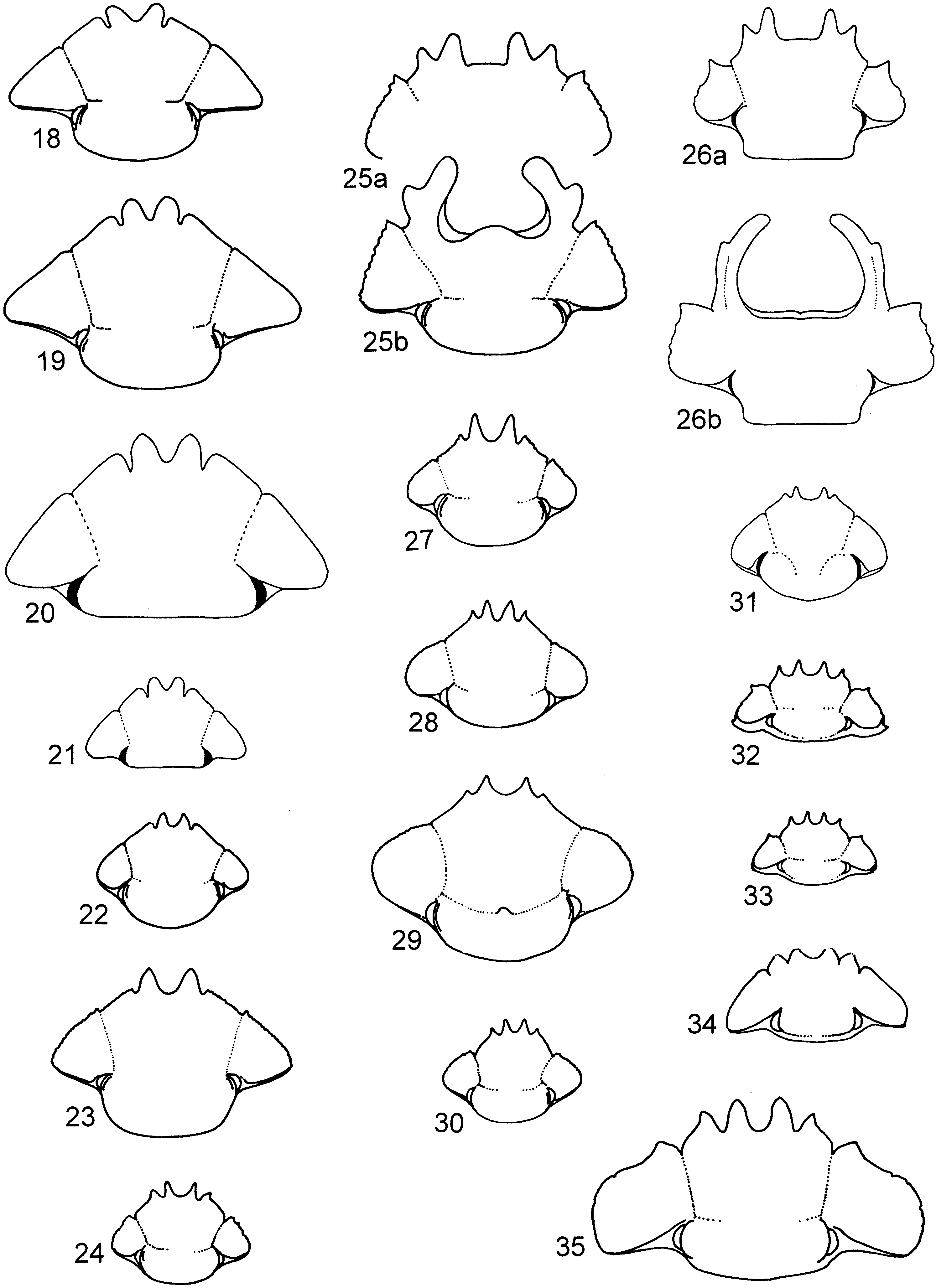
( figures 4 View FIGS , 19 View FIGS , 37, 54, 58)
Pachysoma hippocrates MacLeay, 1821: 507 View in CoL ; MacLeay, 1833: 55; Castelnau 1840: 68; Péringuey, 1902: 77; Gillet, 1911a: 6; Ferreira, 1953a: 16; Ferreira, 1961: 23; Ferreira, 1966: 57; Ferreira, 1969: 21; Holm and Scholtz, 1979; 230. Holotype: no locality (1 3 BMNH).
Pachysoma macleayi Castelnau, 1840: 68 View in CoL : Ferreira, 1961: 23; Holm and Scholtz, 1979: 230.
Pachysoma hessei Ferreira, 1953a: 18 View in CoL ; Ferreira, 1961: 23; Ferreira, 1969: 21; Holm and Scholtz, 1979; 230. Holotype: Namaqualand, Wallekraal (1 3 SAMC).
Scarabaeus hippocrates (MacLeay) : Mostert and Holm, 1982: 275.
Diagnosis. Clypeus bidentate, genae and clypeal edge unserrated and continuous ( figure 19 View FIGS ); protibial spurs bifurcate in both sexes, males have ventral inner edge of protibia elbowed and serrated (this characteristic varies clinally from very marked south of the Olifants River to absent in specimens from the north (i.e. Port Nolloth) ( figures 37a, b); metatarsal claws longer than last tarsal segment; genitalia as in figures 58 a–c.
Distribution, habitat and conservation. Coastal south-western South Africa, recorded from Cape Town, Bloubergstrand (33 ° 48 ∞ S, 18 ° 27 ∞ E) to Port Nolloth (29 ° 15 ∞ S, 16 ° 53 ∞ E) in Namaqualand ( figure 4 View FIGS ). Their habitat preference includes vegetated soft to firm sand of coastal hummocks and hillocks, the periphery of dune systems, and river beds and banks. Scarabeus (Pachysoma) hippocrates occur within the West Coast National Park and the proposed Groen-Spoeg National Park. Habitat modification threatens certain populations of S. (P.) hippocrates , viz. those of Bloubergstrand and Port Nolloth.
Comments on locality data. The Sneeukop (33 ° 07 ∞ S, 19 ° 37 ∞ E) (5000 ft) locality near Wellington, is probably due to an incorrectly labelled specimen. This specimen was supposedly collected by Dr K. H. Barnard, who worked at the SAMC from 1911 to 1964. Dr Barnard was a keen mountaineer, who collected and described 10 species of the high-altitude specialist Colophon Gray (Lucanidae) . Endrödy-Younga (1988) revised Colophon and the only locality label resembling the above is for Colophon stokoei from ‘Upper Snoukop, Wellington, 4500–5000ft., January, K.H.Barnard & R.Primos’. However, a comparison of Barnard’s handwritten locality labels on Colophon specimens in the SAMC with that on the Sneeukop S. (P.) hippocrates specimen reveals that the writing is not Barnard’s, which additionally suggests a mislabelling. No other S. ( Pachysoma ) specimens were collected by Dr Barnard. This single male agrees with S. (P.) hippocrates from Bloubergstrand near Cape Town. Sneeukop is well outside the established coastal distribution range of all S. ( Pachysoma ) species, and no S. ( Pachysoma ) have been reliably collected from high altitudes (1525 m). The Bain’s Kloof pass to Wellington was visited by JduGH and the only remotely suitable habitat for S. ( Pachysoma ) is along the sandy river beds of the Wit and Bobbejaan rivers.
Morphological variation. The S. (P.) hippocrates species complex contains three species, viz. S. (P.) hippocrates , S. (P.) glentoni and S. (P.) endroedyi . It was suspected that the distribution of S. (P.) hippocrates extends from Cape Town to the Olifants River, whereas S. (P.) hessei occurs north of the Olifants River to Port Nolloth. This suspicion was tested, but no distinct change was found in the aedeagal morphology across this potential barrier (Olifants River). Individual aedeagi from disjunct populations, e.g. Port Nolloth (29 ° 15 ∞ S, 16 ° 53 ∞ E) and Modder River (33 ° 28 ∞ S, 18 ° 20 ∞ E) differ significantly, but when all populations were examined there is only evidence of a gradual clinal change. Furthermore, the populations comprising the cline exhibit very slight unique genitalic and other morphological features. This suggests that S. (P.) hippocrates is currently undergoing speciation. Furthermore, within a population the male genitalia can show distinct differences. For example, in the Port Nolloth and Sand Kop (29 ° 42 ∞ S, 17 ° 06 ∞ E) populations the development of a spine on the left paramere is variable. Examination of the external morphology revealed the same trend, but evidently S. (P.) hippocrates sensu lato includes a maincline composed of four subclines (listed below) within it. The main cline (for the entire geographic range of S. (P.) hippocrates ) varies in the following features: (1) aedeagal morphology; (2) variation in size, large in the south to smallest in the north; (3) variation in the expression of the dimorphic elbowed protibia, very slight in the north to very marked in the south. Further detailed examination of all specimens revealed four subclines within S. (P.) hippocrates , i.e. excluding the sister species [ S. (P.) aesculapius , S. (P.) endroedyi and S. (P.) glentoni ]. These populations may be isolated by river courses (i.e. natural barriers) and include: (1) a Cape Town to Lambert’s Bay population, distinguished by large size and dimorphic protibia; (2) an Olifants to Groen River population, recognizable by a reduction in the dimorphism of the protibia, and the presence of waxy indument on the periphery of the elytra; (3) a Groen to Buffels River population, characterized by an even smaller difference in the protibia between sexes, and no waxy indument; (4) a Buffels River to Port Nolloth (possibly extending to the Holgat River, but as yet unconfirmed), characterized by small size, practically no dimorphism of the protibia, but very orange setal colour in mature sclerotized specimens (teneral specimens of other populations may have this colour). Thus, no consistent and easily recognizable suite of characters could be found to justify the existence of S. (P.) hessei at subspecific or specific level. To divide S. (P.) hippocrates into four subspecies would make identification very difficult for a non-specialist. We therefore choose to regard S. (P.) hippocrates as a species with distinct south to north clinal variation, and consequently the synonymy of S. (P.) hessei with S. (P.) hippocrates is maintained.
Biology. Scarabaeus (Pachysoma) hippocrates occur sympatrically with S. (P.) aesculapius south of the Olifants River, and with S. (P.) striatus north of the Olifants River. Of 36 nests excavated by JduGH, five contained only pellets, six contained both detritus and pellets and 25 contained only detritus. This suggests that S. (P.) hippocrates prefers detritus. Two separate attempts were made to breed S. (P.) hippocrates in the laboratory. Three pairs (between 1 September 1996 and 23 January 1997) and six pairs (between 9 September 1998 and 4 December 1998) of S. (P.) hippocrates were placed in round 25 litre buckets and 5 litre square bread bins, respectively. Sand, detritus and dry pellets from their collection site were used. They dug burrows, foraged regularly, but did not breed.
Comments. S. (P.) hippocrates is very similar to S. (P.) glentoni (see diagnosis of these species for the differences) but is easily and reliably differentiated by the male genitalia and geographic distribution.
Types. The holotype of Pachysoma hippocrates has no locality label, but the type locality is recorded by MacLeay (1821) as Cap. Bonae Spei. It is a large (length 35.20 mm, width 22.34 mm) specimen with very worn protibia, and its genitalia were first dissected by JduGH for this study. Holm and Scholtz (1979) suspected it came from the southern extreme of the species’ distribution. The size and male genitalia confirm their suspicion, however, it does not match any labelled material from the southern populations, except a single male labelled C. Bon Spei that conforms to it in size, microsculpture and identical male genitalia.
Ferreira (1953a) listed the holotype, allotype and 12 paratypes of Pachysoma hessei in the SAMC collection, and one paratype in the CPMM collection. Holm and Scholtz (1979) were only able to trace the holotype, allotype and two paratypes in the SAMC collection. The remaining 10 paratypes from the SAMC were discovered during reorganization (1993 onwards) of the collection (see Cochrane, 1995).
Type material examined (S 14 spec. [5], 3 ♀ 11 3, 9 3 diss.). SOUTH AFRICA: HOLOTYPE 3, Pachysoma hippocrates MacLeay : [1] Hippocrates (written on faded white traingle) / 58.60. (written on blue disk) / Type (printed on white disk, surrounded by a red circle) / Pachysoma hippocrates (Kirby MS) , Type McLeay. (written on now faded white paper) / Pachysoma hippocrates M’Lea , Holotype (written on white rectangular card, surrounded by a red line, UPSA label) / Pachysoma hippocrates Holotype dissected J. du G. Harrison 1998 / (1 3 BMNH). HOLOTYPE 3, Pachysoma hessei Ferreira : Wallekraal, Namaqualand, (30.22.05S 17.37.05E) // Mus., Expd., Oct.1950. // (printed locality label) / Holotypus, ♀, Pachysoma Hessei, 1951 , Maria C. Ferreira / Pachysoma hessei Ferreira (= hippocrates M’Leay ), det. Holm & Scholtz 1978 (written on UPSA holotype label) /, (1 3 SAMC), idem, but / Allotypus, ♀, Pachysoma Hessei, 1951 , Maria C. Ferreira /, Pachysoma hessei Ferreira (= hippocrates M’Leay ), det. Holm & Scholtz 1978, (written on UPSA paratype label) /, (1 ♀ SAMC), PARATYPES: idem, but Type, SAM/Ent. 2678, (printed on green card), (8 3 SAMC), (1 3 TMSA), {1x CPMM}; Wallekraal, Namaqualand, (30.22.05S 17.37.05E), S.A.M., Oct.1950 (written) / Type, SAM/Ent. 2678 /, (1 ♀ SAMC), (1 ♀ TMSA).
Additional material examined from South Africa (S308 specs [45], 92 ♀ 185 3, 77 3 diss., 5uns., 26eth., 81p.) .
Scarabaeus (Pachysoma) glentoni Harrison, Scholtz and Chown sp. n. ( figures 5 View FIGS , 20 View FIGS , 38, 55, 59)
Description
Size (mm). Body length, mean 3 29.70; ♀ 29.12; range 3 26.72–36.28; ♀ 25.54–31.98. Pronotal width, mean 3 19.16; ♀ 17.71; range 3 16.38–22.96; ♀ 15.78–18.58 (N= 12 3 12 ♀).
Colour. Cuticle black, setae rufous, antennal clubs golden.
Head ( figure 20 View FIGS ). Dorsal: clypeus bidentate, teeth rounded and separated by V-shaped incision; clypeal and genal margins unserrated, almost continuous, separated at clypeal–genal suture by small incision; whole head finely granular, becoming punctate around periphery; large setiferous granules adjacent eye and genae; posterior edge of genae rounded. Ventral: rim of setae on clypeus and genae edge; clypeal teeth raised into two ridges separated by hollow depression, posterior raised rim with single pointed tooth; line of setae on either side of clypeal ridges; mandibles robust, mentum bilobed, each lobe with stiff setae pointing forwards; antennae lamellate, nine-segmented, club composed of last three segments.
Pronotum. Head amplected into prothorax; anterior lateral edge, irregularly notched, posterior lateral edge, serrate from setal fringe; setal fringe, long on outside edge tapering to short medially; disc globose; midline area unsculptured, centrally widened to form unsculptured diamond-shape; adjacent disc deeply punctate, anterior edges granular; distinct patch of fine granules on posterior lateral edge; posterior edge curved inwards.
Legs. Protibia, dorsal ( figure 38b): quadridentate, teeth pointed, curved downwards, apical surface smooth, basal half transversely furrowed; serrations between teeth; proximal half of protibia with smooth undulations, basal half smooth and continuous; medial setiferous carina; setal border on inner and proximal outer edges, long; setae between teeth, short; inner edge markedly deflected medially; spurs short, stout, bifid and curved downwards. Protibia, ventral ( figure 38a): apical surface of teeth smooth; outer surface scattered with shallow broad punctures, inner edge with fine granules; medial inner lateral edge deflected downwards and bordered with row of irregular but distinct setiferous knobs; medially at base of protibia are two rows, outer composed of small setiferous knobs (about four or five) while inner of carina, both merge into inner protibial deflection. Mesotibia: s hort and robust, outer edge with three bands of seta, inner edge with two bands; mesospur slightly spatulate ( figure 55), apex forms blunt point; tarsal insertion slightly sub-apical; two almost equal in length tarsal claws, both longer than last tarsal segment. Metatibia, dorsal: medial ridge, proximal side smooth, distal side with long setal brush; spurs thin, pointed, round in cross-section, slightly deflected in centre; two equal and curved tarsal claws; claws almost equal to just longer than last tarsal segment. Metatibia, lateral: three curved setal brushes on the tibia.
Elytra. Elytra fused, humeral callus absent; well-developed sub-elytral ridge; elytral rim broad; six faint striae per elytron, most visible when viewed laterally; six interstriae per elytron with rows of irregularly spaced, fine, setiferous granules.
Abdomen. Mesocoxae contiguous, mesosternal ridge absent; abdominal sternites with single row of setiferous granules becoming irregularly spaced as sternite widens laterally; bilobed protuberances in males (absent in females) on lateral edge of sternites 2, 3, 4 (numbered 1–6 from metacoxae) only.
Pygidium. Dorsoventral midline unsculptured; sides scattered with small granules, bilateral smooth rim around edge of pygidium.
Aedeagus. Short, asymmetrical, see figure 59 a–c.
Female. Differs from male as follows; Protibia (ventral). Proximal carina very reduced at (× 40); inner lateral edge with slight deflection, and row of anterior facing setiferous notches (× 40). Abdomen. Width of the last abdominal sternite, broad at pygidium; prominent protuberances on sternites 3–5, absent.
Comments. Scarabaeus (Pachysoma) glentoni is externally virtually indistinguishable from S. (P.) hippocrates . The male genitalia provide the most reliable character for distinguishing these two species ( figure 59 a–c vs 58a–c). Externally S. (P.) glentoni is distinguished from S. (P.) hippocrates by the narrow almost parallel sided mesotarsal spurs ( figure 54 vs 55), and a distinct patch of granules on the posterior of the pronotum. The localized distribution of S. (P.) glentoni ( figure 5a, b View FIGS ) as opposed to the much wider distribution of S. (P.) hippocrates ( figure 4 View FIGS ), separates the two species. Very abraded mesospur of S. (P.) hippocrates ( figure 54 unabraded), resemble the narrow mesospurs of S. (P.) glentoni ( figure 55 unabraded), care should be taken not to confuse the two species on this character.
Distribution, habitat and conservation. Localized to north of Lambert’s Bay and south of the Olifants River, extending inland to Clanwilliam ( figure 5a, b View FIGS ). Known habitats include the firm vegetated sand of river banks (Olifants and Groot- Sandleegte Rivers) and coastal hummocks. S. (P.) glentoni has not been collected in or around larger dunes, and does not occur within any proclaimed conservation area.
Biology. Scarabaeus (Pachysoma) glentoni was observed collecting detritus, which was dragged forward to preconstructed burrows. A single male was observed at its burrow entrance, releasing what was probably pheromone. The beetle adopted a head down position, and using the hind legs, it would rhythmically stroke its abdominal sternites and then flick its legs backwards. This action was interspersed by short breaks (5–10 s), where the hind legs were held outstretched, but not moved. This behaviour was also observed in S. (P.) gariepinus and S. (P.) hippocrates .
Etymology. Named for a keen naturalist, Mr Jon Glenton, who encouraged JduGH’s interest in biology from an early age. Mr Glenton provided the Land-Rover that enabled JduGH to collect this new species.
Type material examined (S124 specs, 35 ♀ 89 3, 18 3 diss., 15 3 eth.). SOUTH AFRICA: HOLOTYPE 3, S.Afr., SW Cape, Nortier Farm , 32.03S- 18.19E / 25.8.1981; E-Y:1847, day, red sand, leg. Endrödy-Younga (1 3 TMSA) GoogleMaps ; idem but GoogleMaps , ALLOTYPE ♀ (1 ♀ TMSA) ; 108 PARATYPES: S.Afr., SW Cape, Nortier Farm , 32.03S- 18.19E / 25.8.1981; E-Y:1847, day, red sand, leg. Endrödy-Younga (4 ♀ 7 3 TMSA), (1 ♀ 1 3 BMNH) GoogleMaps , idem but, SAM-COL-A043134 (1 ♀ 2 3 SAMC) GoogleMaps ; S.Afr., SW Cape, Nortier Farm , 32.03S- 18.19E / 23.8.1981; E-Y:1840, day, red sand, leg. Endrödy-Younga (2 ♀ 5 3 TMSA) GoogleMaps ; S.Afr., SW Cape, Nortier Farm , 32.03S- 18.19E / 25.8.1981; E-Y:1845, groundtraps, leg. Endrödy-Younga / groundtrap with faeces bait (1 3 TMSA) GoogleMaps . S.Afr: Vredendal D., nr. Strandfontein, De Boom 273, Skerpklip , 31 ° 45 ∞ 31 S 18 ° 20 ∞ 55E, 12–14.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG, Harrison 1996, site 199 No .., Grootsandleegte R. Bank, Y / R sand, (4 ♀ 4 3 SANC), (4 3 ZMHB) , idem but, T1425 (5 3 SMWN), (4 3 eth. TMSA) ; S.Afr: Vredendal D., nr. Strandfontein, De Boom 273, Skerpklip , 31 ° 45 ∞ 18 S 18 ° 17 ∞ 38E, 14.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG, Harrison, 1996, site 200, No .., Grootsandleegte R. Bed, white sand, (1 ♀ TMSA) ; S.Afr: Vredendal D., nr. Strandfontein, Byneslaagte 274, Onderputs , 31 ° 47 ∞ 29 S 18 ° 22 ∞ 59E, 14.xii.1996, leg. J.duG. Harrison, site 202, Grootsandleegte R., Y. firm sand plain, (5 3 MNHN), (6 3 BMNH) , idem but, T306 (5 3 BMSA) ; S.Afr: Vredendal D., nr. Strandfontein, Byneslaagte 274, Onderputs , 31 ° 47 ∞ 01 S 18 ° 22 ∞ 11E, 14–15.xii.1996, leg. J.duG. Harrison, site 203, Grootsandleegte R., Y. firm sand plain, (3 ♀ TMSA), (1 3 eth. TMSA) ; S.Afr: Vredendal D., Fonteintjie 466, near Witwater , 31 ° 55 ∞ 17 S 18 ° 22 ∞ 08E, 15.xii.1996, leg. J.duG. Harrison, site 204, sand rise, yellow sand, (5 ♀ 4 3 SANC), (1 3 UPSA), (10 ♀ eth. TMSA) ; S.Afr: Vredendal D., Fonteintjie 466, near Witwater , 31 ° 54 ∞ 58 S 18 ° 21 ∞ 37E, 15.xii.1996, leg. J.duG. Harrison, site 204a, sand rise, yellow sand, (5 3 TMSA), (1 ♀ 2 3 UPSA), (2 ♀ 2 3 COCS) idem but, SAM-COL- A043135 (3 ♀ 5 3 SAMC) , idem but, T1425 (2 ♀ 5 3 SMWN) ; S.Afr: Clanwilliam, nr. Lambert’s Bay, Kookfontein 88, 32 ° 03 ∞ 54 S 18 ° 22 ∞ 39E, 16.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG, Harrison 1996, site 205 No .., Vegt. dune, firm yellow sand (1 ♀ 1 3 TMSA) ; S.Afr. C.P. 12km N., of Clanwilliam , (ca. 32.06S 18.50E), 14 October, 1981, VB Whitehead / SAM-COL-A043133 (1 ♀ SAMC) GoogleMaps ; S. Afr: Clanwilliam , 11.7km W, ca. 32.10S 18.47E, 5.viii.1997, C. R. Owen (3 ♀ 1 3 TMSA); (No locality data) GoogleMaps / Ferreira Collection / NMBH3677 / National Museum of Bloemfontein, Dept. Entomology / 234 (written in blue ballpoint on yellow card, possibly by Ferreira ) / T306 (1 3 BMSA) .
Scarabaeus (Pachysoma) endroedyi Harrison, Scholtz and Chown sp. n. ( figures 5 View FIGS , 21 View FIGS , 39, 56, 60)
Description
Size (mm). Body length: mean 3 22.92; ♀ 24.28; range 3 20.66–24.70; ♀ 22.30–26.40. Pronotal width: mean 3 14.54; ♀ 14.20; range 3 12.78–15.98; ♀ 12.84–15.18 (N= 12 3 12 ♀).
Colour. Cuticle black; elytral indument grey-white to brown; antennal club golden; seta rubescent to brown.
Head ( figure 21 View FIGS ). Dorsal: clypeus bidentate; teeth short, apex rounded, separated by smooth V-shaped incision; genal and clypeal edge unserrated, almost continuous, separated at genal–clypeal incision; head finely granular, setose granules restricted to genae and posterior of head between eyes. Ventral: rim of setae around clypeal and genal edge; clypeal teeth raised into two ridges, posterior rim, with single pointed tooth; mandibles robust, mentum bilobed, stiff setae pointing forward on lobe; lamellate antennae, nine-segmented, club formed by last three segments, which are covered with fine golden setae; ventral eye smaller than dorsal eye.
Pronotum. Head amplected into prothorax; concavity bordered by bilateral rim; anterior lateral edge, unequally notched; posterior lateral edge, serrate from setal fringe; fringe, long on outer edge tapering to short medially; disc globose and setiferously granula-punctate; medial diamond-shape and posterior lateral edges, finely shagreened.
Legs. Protibia, dorsal ( figure 39b): quadridentate, teeth pointed, curved downward, apical surface smooth, basal half transversely furrowed; blunt serrations between and proximal to teeth, becoming smooth on proximal half of protibia; medial setiferous carina; setal border, long on inner edge and proximal outer edge, short between teeth; inner edge, deflected inwards medially; spur weakly bifid and curved downward. Protibia, v entral ( figure 39a): surface granular, apical section of teeth smooth; proximal carina and single row of setae dissipate before proximal tooth; inner edge deflected downwards, bordered with row of irregular, distinct knobs. Mesotibia: mesotibia short and robust; outer edge with three bands of seta; spurs slightly spatulate, apex blunt; tarsal insertion slightly sub-apical; two unequal tarsal claws, both shorter than last tarsal segment. Metatibia, dorsal: proximal side of medial ridge smooth, distal side with long setal brush; spurs thin, apex pointed, round in cross-section, slightly deflected in centre; two equal and curved tarsal claws, shorter than or almost equal to last tarsal segment. Metatibia, lateral: three curved setal brushes on tibia.
Elytra. Fused, humeral callus absent; well-developed sub-elytral ridge and rim; elytra bordered by band of indument; six striae per elytron, scattered with minute, irregular setose granules.
Abdomen. Mesocoxae contiguous, mesosternal ridge absent; abdominal sternites with single row of setiferous granules becoming irregularly spaced as sternite widens laterally; bilobed protuberances in males (absent in females) on lateral edge of sternites 2, 3, 4 (numbered 1–6 from metacoxae) only.
Pygidium. Irregularly speckled with small granules; bilateral smooth rim around edge of pygidium.
Aedeagus. Parameres short, asymmetrical, see figure 60 a–c.
Female. Differs from male as follows: Protibia, ventral: proximal carina and single row of setae very reduced at (× 40); inner lateral edge with slight deflection, and row of anterior facing setiferous notches (× 40). Abdomen: width of the last abdominal sternite, broad at pygidium; prominent protuberances on sternites 3–5 wanting.
Comments. The smallest species within the hippocrates species complex. Similar to S. (P.) hippocrates but with elytral indument like S. (P.) striatus and S. (P.) gariepinus . The sympatric population of S. (P.) hippocrates also exhibits elytral indument, but to a lesser degree. The two species are easily distinguished as S. (P.) hippocrates is larger and has metatarsal claws longer than the last tarsal segment, while S. (P.) endroedyi metatarsal claws are shorter.
Distribution, habitat and conservation. Restricted to south-western South Africa, occurring just north of the Olifants River. The localized distribution ( figure 5a, b View FIGS ) might be an artifact of the absence of roads passing through their habitat (thus preventing easy collection), or lack of suitable habitats further northwards. Preferred habitats include the vegetated firm sand of coastal hillocks and sand dunes. This species does not occur within a proclaimed conservation area. The increase in recreational development around Strandfontein (31 ° 45 ∞ S, 18 ° 14 ∞ E), and Koekenaap (31 ° 32 ∞ S, 18 ° 14 ∞ E) potentially threatens the future survival of this species.
Biology. Seven nests of this species were excavated by JduGH. One contained pellets and six contained a combination of detritus and pellets. S. (P.) endroedyi thus may be a mixed dung and detritus feeder. All specimens were collected during daylight. Their foraging behaviour is typical of S. ( Pachysoma ). The larvae are unknown.
Etymology. Named for the late Dr Sebastian Endrödy-Younga, who recognized this species as new and collected most of the type series. JduGH is indebted to Sebastian for encouraging and guiding his interest in beetle collecting and systematics.
Type material examined (S135 specs [0], 47 ♀ 88 3, 16 3 diss.). SOUTH AFRICA: HOLOTYPE 3, S.Afr: Namaqualand, Kommandokraal farm, 31.30S- 18.12E / 23.9.1994; E-Y:3033, on sandy ground, Endrödy & Bellamy, (1 3 TMSA) GoogleMaps ; idem, but GoogleMaps ALLOTYPE ♀, (1 ♀ TMSA) ; idem, but PARATYPES, (2 ♀ 4 3 BMNH), (12 ♀ 23 3 TMSA), (2 3 UPSA), (2 3 MNHN), (2 ♀ ZMHB) , idem, but T307 (2 3 BMSA) , idem, but T1424 (2 3 SMWN) ; S. Afr., Namaqualand, Kommandokraal Frm. , 31.30S- 18.12E / 30.8.1979; E-Y:1622, singled on sand, leg. Endrödy-Youngae, (2 3 TMSA) GoogleMaps ; S.Afr., Namaqualand, Koekenaap , 31.32S- 18.14E / 30.8.1979; E-Y:1623, red dunes night, leg. Endrödy-Youngae, (2 3 TMSA) GoogleMaps ; S. Afr., Namaqualand, Koekenaap , 31.32S- 18.14E / 31.8.1979; E-Y:1624, red dunes day, leg. Endrödy-Youngae, (4 ♀ 5 3 TMSA) GoogleMaps ; S.Afr., Namaqualand, Koekenaap , 31.32S- 18.14E / 31.8.1979; E-Y:1625, groundtraps, 59 days, leg. Endrödy-Youngae / groundtraps with banana bait (1 3 TMSA) GoogleMaps , idem, but groundtraps with meat bait (1 3 TMSA) GoogleMaps ; idem, but groundtraps with faeces bait (1 3 TMSA) GoogleMaps ; South Africa: Cape, Kommandokraal , 31.31S- 18.13E, 100m, 23-IX-1985, AVEvans, CLBellamy / T1424 (2 ♀ 4 3 SMWN) GoogleMaps ; S. Africa, C.P., Kommandokraal 624, Vredendal , 31 ° 30 ∞ S, 18 ° 12 ∞ E / 23 Sept. 1985, Louw, v. Rensburg, NMBH 15818 / T307 (1 ♀ BMSA) ; Kommandokraal , Cape, R. S.A., 31 ° 30 ∞ S,. 18 ° 12 ∞ E., 19 Sept. 1982, M.-L. Penrith / H54679 / T1424 (1 ♀ SMWN) ; S.Afr.; W Cape, KommandokraalfarmW (sic), 31.29S- 18.11E / 23.9.1994; E-Y:3035, on ground, Endrödy & Bellamy (5 3 TMSA) GoogleMaps ; S.Afr; Namaqualand, Koekenaaphilldunes (sic), 31.32S- 18.14E / 23.9.1994; E-Y:3032, groundtraps, 3 days, Endrödy & Bellamy / groundtraps with banana bait (1 3 TMSA) GoogleMaps ; S.Afr; SW Cape, Koekenaap dunes, 31.32S 18.14E / 22.9.1994; E-Y:3030, ground & hummocks, Endrödy & Bellamy (6 ♀ 3 3 TMSA) GoogleMaps , idem, but SAM-COL-A043137 (6 ♀ 3 3 SAMC) GoogleMaps ; S.Afr: Vredendal D., Graafwater on Farm 156, 31 ° 22 ∞ 38 S 18 ° 01 ∞ 23E, 6.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG. Harrison 1996, Site 186 No .., firm Y. sand plain, nr. veget. dunes (1 3 TMSA) ; S. Afr : Vredendal D., Elsie Erasmus Kloof 158, 31 ° 24 ∞ 46 S 18 ° 02 ∞ 30E, 7–8.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG. Harrison, 1996, site 188 No .., vegt. ‘old’ dune, yellow sand, (2 ♀ 4 3 BMNH) , idem, but T1424 (2 ♀ 4 3 SMWN) ; S.Afr: Vredendal D., Kommandokraal, on Farm 624, near Koekenaap , 31 ° 29 ∞ 58 S 18 ° 11 ∞ 47E, 8–9.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG. Harrison 1996, site 191 No .., vegetated dune, yellow sand, (1 3 SANC) ; S.Afr: Vredendal D., Kommandokraal, on Farm 624, near Koekenaap , 31 ° 29 ∞ 02 S 18 ° 10 ∞ 08E, 10.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG. Harrison 1996, site 193 No .., vegetated dune, yellow sand, (1 ♀ 2 3 SANC) ; S.Afr: Vredendal D., Vleitjies, Farm 620 near Koekenaap , 31 ° 34 ∞ 53 S 18 ° 10 ∞ 15E, 11.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG. Harrison 1996, site 195 No .., Olifants R. vegt., dune/bank, red sand, (2 ♀ 5 3 SANC) ; S.Afr: Vredendal D., Vleitjies, Farm 620 near Koekenaap , 31 ° 34 ∞ 53 S 18 ° 10 ∞ 56E, 12.xii.1996, leg. J.duG. Harrison / Univ.Pret.Zoo & Ento., J. duG. Harrison 1996, site 196 No .., Olifants R., vegt. dune, soft red sand / T307 (2 ♀ 1 3 BMSA) .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Scarabaeus (Pachysoma) hippocrates ( MacLeay, 1821 )
| Harrison, J. Du G., Scholtz, C. H. & Chown, S. L. 2003 |
Scarabaeus hippocrates (MacLeay)
| MOSTERT, L. E. & HOLM, E. 1982: 275 |
Pachysoma hessei
| FERREIRA, M. C. 1961: 23 |
| FERREIRA, M. C. 1953: 18 |
Pachysoma macleayi
| HOLM, E. & SCHOLTZ, C. H. 1979: 230 |
| FERREIRA, M. C. 1961: 23 |
| CASTELNAU, L. 1840: 68 |
Pachysoma hippocrates
| FERREIRA, M. C. 1966: 57 |
| FERREIRA, M. C. 1961: 23 |
| FERREIRA, M. C. 1953: 16 |
| GILLET, J. J. E. 1911: 6 |
| CASTELNAU, L. 1840: 68 |
| MACLEAY, W. S. 1833: 55 |
| MACLEAY, W. S. 1821: 507 |