
Scalenodon, MONOPHYLY AND THE MANDA BEDS
|
publication ID |
https://doi.org/10.1111/j.1096-3642.2003.00096.x |
|
persistent identifier |
https://treatment.plazi.org/id/03EE87B5-9745-1548-FCE2-E515FE65FB26 |
|
treatment provided by |
Carolina |
|
scientific name |
Scalenodon |
| status |
|
SCALENODON MONOPHYLY AND THE MANDA BEDS
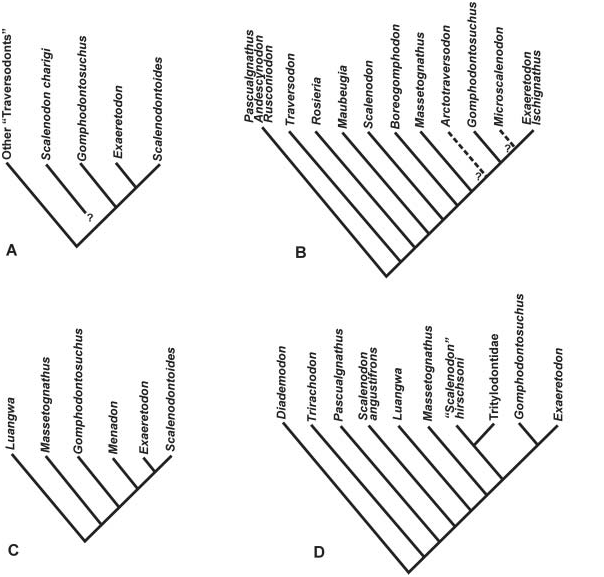
It is interesting to compare the different ‘species’ of Scalenodon from the Manda Formation in Tanzania. Crompton (1955) initially described S. angustifrons from material previously proposed as Trirachodon angustifrons by Parrington (1946). Crompton (1972) ( Figs 7 View Figure 7 , 10A- C View Figure 10 ) recognized three additional species from the same beds: S. hirschsoni , S. attridgei and S. charigi . Considering the wide morphological variation between these taxa, Hopson (1984) suggested that they could be separated at the generic level, and later ( Hopson, 1985) proposed a possible closer relationship between S. charigi with a clade comprising Gomphodontosuchus , Exaeretodon , and Scalenodontoides ( Fig. 7A View Figure 7 ).
More recently, the phylogenetic proposal of Hopson & Kitching (2001, using a parsimony program) found that S. angustifrons and ‘ S. ’ hirschsoni were distant relatives (i.e. the genus Scalenodon was polyphyletic; Fig. 7D View Figure 7 ). Even in a different context, considering that they proposed a sister-group relationship of ‘ S. ’ hirschsoni with tritylodontids, our phylogeny confirms the distant relationship between S. angustifrons and ‘ S.’ hirschsoni . In addition, we agree with Hopson (1984) that the other two species included in Scalenodon should be placed in different genera, given the remarkable differences in their postcanine morphology (although ‘ S.’ attridgei and ‘ S.’ charigi may prove to be the same taxon).
The Manda fauna shows an unusual mixture of clearly plesiomorphic forms (e.g. S. angustifrons ) and ‘ S. ’ hirschsoni and ‘ S. ’ charigi , which share derived postcanine features with late Ladinian-Carnian forms ( Hopson, 1984). This kind of assortment, unknown for traversodontid faunas until recently, was also reported from Madagascar ( Flynn et al., 1999, 2000) and Brazil ( Abdala et al., 2001). In both cases, taxa morphologically closer to Ladinian traversodontids, and others more closely related with typical Carnian ones, were found in association. The condition in the Manda Formation is more extreme because the fauna includes Diademodon -like forms and Cricodon , closely related to Trirachodon , suggesting a late Scythian/ early Anisian age ( Kitching, 1995; Hancox, 2000), whereas traversodontids like ‘ S. ’ hirschsoni and ‘ S. ’ charigi have features known otherwise in Ladinian and Carnian traversodontids. S. angustifrons , both for its morphological features, and by its direct association with the rhynchosaur Stenaulorhynchus and cynodont mandibles with diademodontid-like teeth ( Crompton, 1955), seems to be older, whereas the other species of ‘ Scalenodon ’ were seemingly isolated occurrences from different outcrops ( Crompton, 1972). The extreme differentiation between these traversodontids suggests that, as proposed by Boonstra (1953) and, on the basis of more extensive evidence, by Charig (1963), more than one age could be represented in the fauna of the Manda Formation (see also Gay & Cruickshank, 1999: 199).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.