
Salka diversa, Ohara, Naomichi, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.280852 |
|
DOI |
https://doi.org/10.5281/zenodo.6181872 |
|
persistent identifier |
https://treatment.plazi.org/id/03CF2470-9614-4709-6380-F98BFA9C3024 |
|
treatment provided by |
Plazi |
|
scientific name |
Salka diversa |
| status |
sp. nov. |
Salka diversa View in CoL sp. nov.
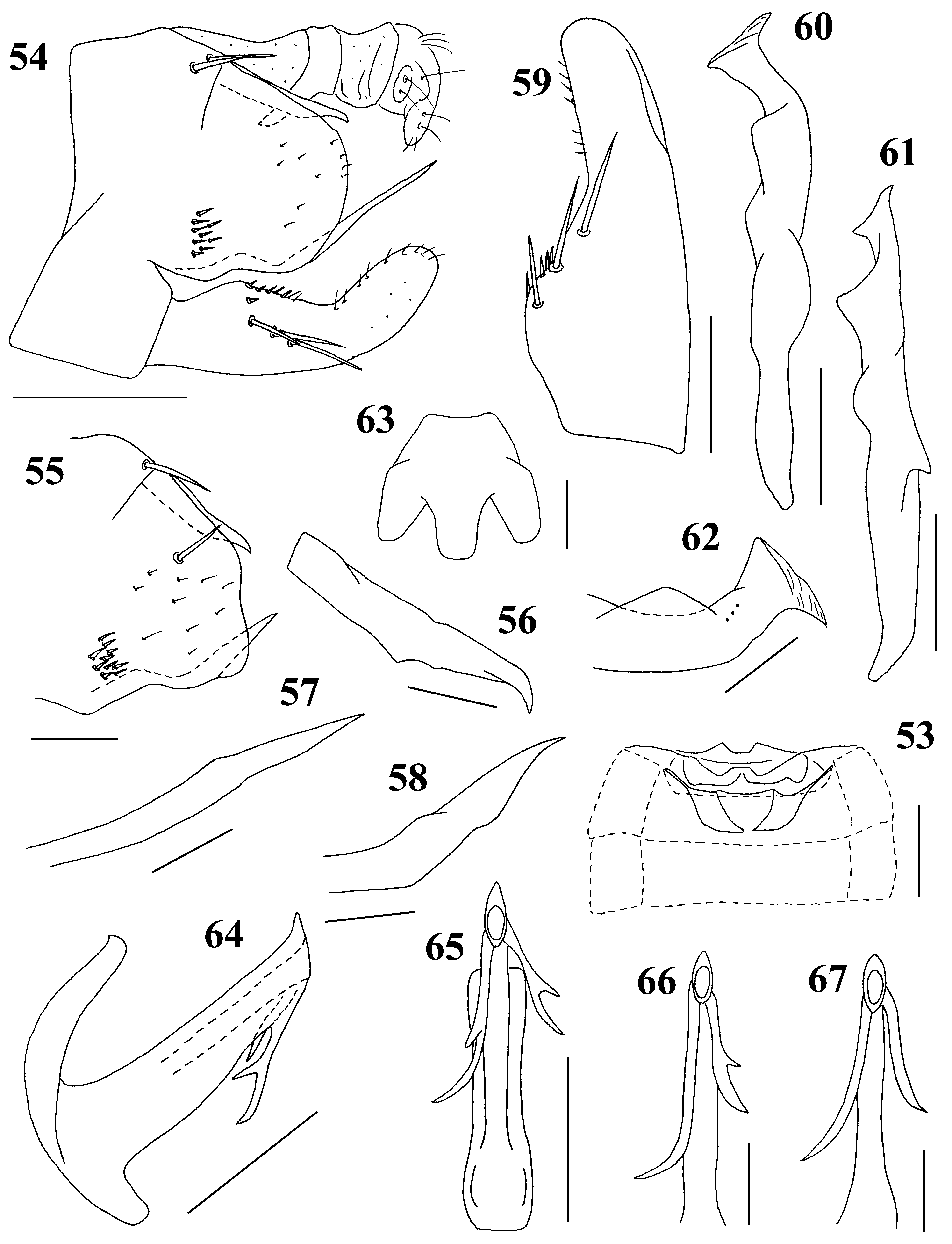
( Figs. 4 View FIGURES 1 – 8 , 12 View FIGURES 9 – 16 , 53–67 View FIGURES 53 – 67 , 116 View FIGURES 113 – 121 )
Male body brown, infuscated in some paratypes. Female often paler and yellowish, sometimes similar to male. Vertex pale yellow, bearing black spot centrally. Pronotum with anterior margin pale yellow; mesonotum pale yellow, with basal triangles brown indistinct; fore wing with brochosome field infuscated. Abdomen infuscated; male subgenital plates pale brown to brown; ovipositor (3rd valvulae) ochreous to brown.
Vertex 2.0 times as wide as long at dorsal mid-line; coronal suture distinct basally. Pronotum 1.8 times as wide as long; mesonotum slightly shorter than pronotum. Male abdominal sternal apodemes not reaching posterior margin of 3rd sternite. Female 7th abdominal sternite pentagonal, with posterior margin swollen. Ovipositor (3rd valvulae) obviously extending beyond pygofer.
Body length (mean): 3, 2.9–3.3 mm (3.1 mm); Ƥ, 2.9–3.4 mm (3.2 mm).
Male genitalia ( Figs. 54–67 View FIGURES 53 – 67 ). Pygofer rounded or slightly sinuate caudally, bearing 1–3 macrosetae on dorsal margin, tuft of short macrosetae at lower basal angle, numerous short setae scattered and rarely macrosetae on disc of lobe, with dorsal and ventral processes; dorsal process slightly exceeding pygofer, weakly curved ventrad near apex; ventral process thin, sinuate near base, tapering, extending well beyond pygofer ( Figs. 54, 57 View FIGURES 53 – 67 ; holotype) or sinuate, thick, tapering, slightly extending beyond pygofer ( Figs. 55, 58 View FIGURES 53 – 67 ; some paratypes). Subgenital plate widened in basal 2/3, bearing 3–5 macrosetae. Style almost uniform in width; apophysis of style long, ca. 0.2 times as long as style with some minute furrows apically. Connective U-shaped, with central lobe distinct. Aedeagus with dorsal apodeme elongate; shaft nearly straight bearing pair of asymmetrical apical processes; each apical process usually with small projection at middle of lateral margin ( Fig. 65 View FIGURES 53 – 67 ; holotype) or occasionally without projection ( Figs. 66, 67 View FIGURES 53 – 67 ; some paratypes); preatrium short; gonopore apical on caudal surface.
Type series. Holotype: 3, Mt. Nesugata-yama, Amagi/Tokunoshima, Tokunoshima Is., Ryukyus, Japan, 4. VII. 2007, M. Hayashi leg. Paratypes: [Amami-Oshima Is.] 13, Mt. Matsunaga-yama, Amami-Naze, 19. IX. 2006, M. Hayashi et al.; [Tokunoshima Is.] 13, same data as holotype; 23 1Ƥ, Mt. Inokawa-dake, Tokunoshima, 2. VII. 2007, M. Hayashi leg.; 13, same data except 5. VII. 2007; [Okinawa Is.] 1Ƥ, Mt. Terukubi-yama, Kunigami, 22. X. 1990, M. Hayashi et al.; 13, same data except 5. V. 1991; 23 1Ƥ, same data except 23. XII. 1991; 1Ƥ, same data except 16. V. 1993; 133 21Ƥ, same data except 21.II. 2008; 1Ƥ, same data except 2. III. 2009; 133 13Ƥ, same data except 12. XII. 2009; 23 1Ƥ, Mt. Yonaha-dake, Kunigami, 24. III. 1993, M. Hayashi et al.; 63 6Ƥ, same data except 8. X. 1995; 1Ƥ, same data except 2. VII. 1996; 1Ƥ, same data except 3. IV. 1999; 33 7Ƥ, same data except 12. IX. 2005; 23 5Ƥ, same data except 22. III. 2010, N. Ohara leg. ( ELKU); 13 1Ƥ, Mt. Nishime-dake, Kunigami, 16. V. 1993, M. Hayashi et al.; 1Ƥ, same data except 18. VI. 1994; 1Ƥ, same data except 21. II. 2008; 33 1Ƥ, same data except 22. III. 2010, N. Ohara leg. ( ELKU); 3Ƥ, Oku For. Rd., Kunigami, 2. VII. 1996, M. Hayashi et al.; 13, same data except 12. IX. 2005; 33 1Ƥ, same data except 29. IX. 2007; 63 4Ƥ, same data except 2. III. 2009; 1Ƥ, Ibu, Kunigami, 21. II. 2008, M. Hayashi et al.; 33 1Ƥ, Mt. Fenchiji-dake, Kunigami, 22. III. 2010, N. Ohara leg. ( ELKU); 1Ƥ, Okuma, Kunigami, 11. IX. 2005, M. Hayashi et al.; 13, Aha, Kunigami, 21. II. 2008, M. Hayashi et al.; 13, Hedo, Kunigami, 16. V. 1993, M. Hayashi et al.; 13, Henan, Ôgimi, 11. XII. 2009, M. Hayashi et al.; 1Ƥ, Shioya, Ôgimi, 6. IV. 1991, M. Hayashi et al.; 13 1Ƥ, Mt. Nekumachiji, Ôgimi, 28. IX. 2007, M. Hayashi et al.; 2Ƥ, Kawata, Higashi, 1. III. 2009, M. Hayashi et al.; 73 2Ƥ, Genka-Ôkawa, Nago, 28. IX. 2007, M. Hayashi et al.; 163 10Ƥ, same data except 20. II. 2008; 13 2Ƥ, same data except 11. XII. 2009; 1Ƥ, Kyoda, Nago, 30. IV. 1991 (light trap), M. Hayashi et al.; 13, Ôshittai, Nago, 19. III. 2010, N. Ohara leg. ( ELKU); 4Ƥ, Mt. Katsuu-dake, Nago, 9. X. 1995, M. Hayashi et al.; 13, same data except 22. II. 2008; 13, Nago Central Park, Nago, 14. IX. 2005, M. Hayashi et al.; 13, Noborimata, Nakagusuku, 23. II. 2008, M. Hayashi et al.; 1Ƥ, Kisembaru, Kin, 4. VII. 1996, M. Hayashi et al.; 1Ƥ, Yamada, Onna, 8. IV. 1991, M. Hayashi et al. The holotype is deposited in the Department of Biology, Faculty of Education, Saitama University, Saitama, Japan.
Distribution. Japan (Ryukyus: Amami-Oshima Is., Tokunoshima Is., Okinawa Is.).
Remarks. This species shows some geographical variations in the configuration of the male genitalia. The ventral pygofer process of specimens collected from Amami-Oshima Is. and Tokunoshima Is. ( Figs. 54, 57 View FIGURES 53 – 67 ) is thinner and longer. Furthermore, a pair of aedeagal apical processes is highly varied in the material from Okinawa Is.; often both apical processes have a small projection, occasionally one or rarely both apical processes have no accessory projection. This species is similar to S. jiangshiensis Zhang, Yang et Huang., 2009 described from southern China (Fujian), but can be distinguished by the male genitalia: aedeagus with shaft straight, provided with apical process (without ventral process).
Bionomics. This species probably inhabits the herbaceous layer along woodland paths or shady peripheries of subtropical forests, but the host plants are unknown. Adults seem to occur throughout the year.
Etymology. The specific name is derived from the variations in the configuration of male genitalia.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Typhlocybinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Typhlocybinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Typhlocybinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Typhlocybinae |
|
Genus |