Amphibia
View in CoL
and Reptilia
The Volchaya Balka and Gaverdovsky localities yielded 14 taxa of amphibians and 12 taxa of reptiles:
Mioproteus caucasicus ESTES et DAREVSKY, 1977 (Proteidae)
,
Chelotriton paradoxus POMEL, 1853
,
Triturus cristatus (LAURENTI, 1768)
,
Triturus cf. marmoratus (LATREILLE, 1800)
,
Lissotriton sp. (Salamandridae)
,
Latonia sp. (Alytidae)
,
Bombina cf. bombina (LINNAEUS, 1761) (Bombinatoridae)
,
Hyla sp. (Hylidae)
,
Palaeobatrachus sp. (Palaeobatrachidae)
,
Pelobates sp. (Pelobatidae)
,
Bufotes viridis (LAURENTI, 1768)
,
Bufo sp. (Bufonidae)
,
Rana sp.
,
Pelophylax sp. (Ranidae)
,
Emys cf. tarashchuki (CHKHIKVADZE, 1980) (Emydidae)
, cf.
Sakya sp. (Geoemydidae)
,
Testudinidae
indet.,
Testudo
s. s. (
Testudinidae
),
Pseudopus pannonicus (KORMOS, 1911)
,
Anguis sp. (Anguidae)
,
Lacertidae
indet. (
Lacertidae
),
Coronella sp.
,
Natrix
cf. longivertebrata SZYNDLAR, 1984,
Natrix sp.
, Colubrinae indet. (
Colubridae
), and
Viperidae
indet. (
Viperidae
).
Amphibia LINNAEUS, 1758
The Amphibia
View in CoL
are represented by the
Anura
View in CoL
and Urodela. The tailed amphibians (Urodela) are represented by relatively large forms such as
Mioproteus
and
Chelotriton
and several smaller forms of
Triturus
View in CoL
sensu lato. Remains of
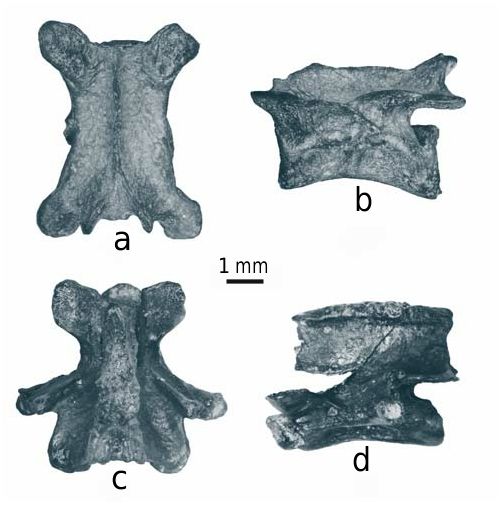
Mioproteus caucasicus
are relatively rare and represented by eleven vertebrae ( Text-fig. 7a, b
View Text-fig
). Their overall morphology is consistent with
M. caucasicus
described from the same area (Maikop city park), but from slightly older deposits, in the shape of the forking processes on the dorsoposterior region of the neural arch and other characters ( Estes and Darevsky 1977).
Chelotriton
is more frequent than
Mioproteus
and documented by about 40 cranial and postcranial (vertebrae) elements ( Text-fig. 7c, d
View Text-fig
). It is assigned to
Ch. paradoxus POMEL, 1853
(according to Schoch et al. 2015) based on the heavily ossified skull bones and extensive tubercular ornamentation.
Triturus
View in CoL
sensu lato is the most abundant tailed amphibian in Volchaya Balka and Gaverdovsky (several hundred vertebrae and skeletal elements) and represented by at least three forms. These are relatively large size
Triturus cristatus
View in CoL
(Pl. 7, Fig. 1a, b) and
Triturus cf. marmoratus
View in CoL
(Pl. 7, Fig. 2a, b) which differ in the length of the neural arch (long in
T. cristatus
View in CoL
and short in
T. cf. marmoratus
View in CoL
) and height of the neural spine (low in
T. cristatus
View in CoL
and high in
T. cf. marmoratus
View in CoL
), and smaller
Lissotriton sp.
(Pl. 7, Fig. 3a, b) with a very high neural spine. Additionally, two fragmentary opisthocoelous vertebrae which are different from
Chelotriton
in the absence of ornamentation and from
Triturus
View in CoL
s. str. and
Lissotriton
View in CoL
in its relatively large size and flattened posterior part of neural arch and, thus, indicate the presence of another salamandrid taxon.
Bombina
View in CoL
is very rare in the material and represented by seven elements. It is indicated by the opisthocoelous presacral vertebrae, posterior position of tuber superior of the ilia and poorly developed pars descendens (Pl. 7, Figs 4, 5). The Caucasian
Bombina
View in CoL
shows the presence of a prominent tuber superior and preacetabular fossa which is characteristic of
B. bombina
View in CoL
rather than
B. variegata
View in CoL
( Böhme 1977, Sanchíz and Młynarski 1979). One of the most common anurans was
Latonia
View in CoL
which is represented by 70 cranial and postcranial elements (Pl. 7, Fig. 6). It differs from
Latonia gigantea (LARTET, 1851)
, the most widespread species in the Late Miocene of Europe ( Rage and Roček 2003), in the maxilla lacking any sculpture. Caucasian
Latonia
View in CoL
also differs from other existing
Latonia species
in shape of sacral transverse processes and possibly belongs to a separate species.
Hyla
View in CoL
is extremely rare and represented by four ilia and a single sacral vertebra (Pl. 7, Fig. 7). Its presence is confirmed by the morphology of the ilia, i.e., thin and expanded anteriorly anteroventral margin of the pars descendens. The tuber superioris is prominent and projects anteriorly. It is nearly oval in shape, which allows distinguishing between
Hyla arborea
View in CoL
(with an oval tuber superior) and
H. meridionalis
View in CoL
(with a rounded tuber superior) ( Holman 1992, Bailon 2000). Because the osteological characters of
Hyla savignyi
View in CoL
are not known, the Caucasian
Hyla
View in CoL
can only be identified to the genus. Another common anuran is
Palaeobatrachus
which is represented by 50 cranial and postcranial elements (Pl. 7, Figs 8, 9). Among the other palaeobatrachids it clearly differs from the older (Oligocene – Early Miocene)
Palaeobatrachus species
and is most similar to the Plio-Pleistocene species (
P. eurydices
and
P. langhae
) in the reduction of premaxillar and maxillar teeth. The
Pelobatidae
View in CoL
, which are predominantly fossorial (burrowing) frogs are fairly rare here (about 15 cranial bones, vertebrae and ilia; Pl. 7, Fig. 10). Most available remains were assigned to the extant genus
Pelobates
View in CoL
. As in the other
Pelobates
View in CoL
, it is characterized by missing dorsal crests on the iliac shafts and striated scars on the postero-mesial border of the ilia. As in the Miocene
Pelobates
View in CoL
it has a small interiliac tubercle ( Rage and Hossini 2000). In addition to
Pelobates
View in CoL
, the material from Volchaya Balka also includes several remains of the fossil genus
Eopelobates
with characterictic pit-and-ridge sculpture on frontoparietals ( Syromyatnikova 2017).
Bufonidae
View in CoL
are documented by toothless premaxilla and lateral position of the olecranon scar on the humerus and several fragmentary ilia. The presence of small bulges on the lateral side of the tuber superior on the ilium and preacetabular fossa indicates the toad species
Bufotes viridis
View in CoL
(Pl. 7, Fig. 11). In the material it is represented by the less than 10 bones. The single humerus which is assigned here to
Bufo sp.
can be distinguished from the former species by its larger size (about twice as big as
B. viridis
View in CoL
).
Ranidae
View in CoL
are the most numerous in the amphibian material and are represented by more than 300 cranial and postcranial elements (Pl. 7, Figs 12, 13). Some specimens exhibit characters typical for brown frogs
Rana
View in CoL
(
Rana temporaria
View in CoL
group), i.e., lateral position of the parietal eminence of the frontoparietal, dorsally inflected medial and lateral crests of humerus, presence of tubercles on tuber superior of the ilia ( Böhme 1977, Ratnikov 2001). Most of the ranid bones exhibit characters typical for green frogs
Pelophylax
View in CoL
(
Rana esculenta
View in CoL
group) (Pl. 7, Fig. 14), i.e., elongate external mandibular ridge of angular, extremely expanded and flattened pars epicoracoidalis and slender middle part of coracoid, high dorsal crest (much higher than iliac shaft) of ilium, laterally flattened tuber superior and its position close to the front of the anterior edge of the acetabulum ( Böhme 1977, Böhme and Günther 1979, Rage 1984, Ratnikov 2001). Extant ranid species are mainly distinguished by external morphological characters. Thus, the described material of
Rana sp.
and
Pelophylax sp.
is not assigned here to extant species.
Reptilia MCCARTNEY, 1802
Turtles are not numerous in the studied localities and are preserved as isolated shell plates. Six species are present belonging to the three families:
Emydidae
,
Geoemydidae
and
Testudinidae
. The majority of turtle specimens belong to the emydids. Some are similar to
Emys tarashchuki
(described from the Late Miocene of Ukraine) in the position of skinscalesulcusontheepiplastron, whichisrelativelydistantfrom the free edge of the bone.
Geoemydidae
are represented by
Sakya sp.
, which was identified based on the peculiar shield configurations, i.e., having an extra number of vertebral, pleural and marginal scutes on the carapace ( Chkhikvadze 1983).
Testudinidae
are represented by a fragment of the anterior lobe of the plastron (epi+entoplastron) from the Gaverdovsky locality and several isolated shell plates. Some fragments of xiphiplastra indicate the presence of a hypoxiphiplastral hinge and can be referred to
Testudo
s. s.
Remains of squamates document the presence of at least six taxa. The anguids are represented, first of all, by
Pseudopus pannonicus
(Pl. 7, Fig. 15), the species common in the European Late Miocene. The remains of this lizard are abundant and include
a posterior
part of the braincase, i.e., a diagnostic element within the genus, vertebrae and osteoderms.
Anguis
are slightly more frequent than
Pseudopus
and recognized on the basis of the frontal morphology, presence of curved and widely-spaced teeth and non-rectangular osteoderms without keels (Pl. 7, Figs 16, 17). Lacertids are represented by a limited quantity of small fragments of lower jaws and vertebrae which does not permit an accurate identification of genus and species (Pl. 7, Fig. 18).
Coronella sp.
is represented by several small size vertebrae with a dorso-ventrally flattened neural arch and weakly developed haemal keel (Pl. 7, Fig. 19). The Caucasian
Coronella
is most similar to
C. miocaenica VENCZEL, 1998
(Middle Miocene, MN 8, Romania and Late Miocene, MN 13, Hungary) in its less depressed neural arch ( Venczel 1998), however its centrum length/width ratio is 1.44, which corresponds to
C. austriaca
rather than
C. miocaenica ( Szyndlar 1991a)
. Three fragmentary vertebrae document the presence of another colubrine which differs from
Coronella
in a larger size and vaulted neural arch. The four fragmentary vertebrae are assigned to
Natrix
cf. longivertebrata, a widely distributed species in the European Neogene ( Szyndlar 1991b), based on their elongate and narrow centra with strong subcentral ridges. Another taxon of
Natrix
is characterized by its relatively small size and centrum length/centrum width ratio of 1.38 (Pl. 7, Fig. 20). It is most similar to
N. rudabanyaensis SZYNDLAR, 2005
(Middle – Late Miocene, MN 6–9, Hungary and Romania) in its moderately developed subcentral ridges, centrum length/ width ratio and, rounded distal hypapophyseal tip ( Szyndlar 2005). The single fragmentary trunk vertebra is assigned to the
Viperidae
, however, due to the limited amount of the material, the identification is possible to the family rank only.