
Rhyacodrilus longichaeta, Rodriguez, Pilar & Fend, Steven V., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3664.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C8136C89-7787-477D-BC64-97AA6C16057B |
|
DOI |
https://doi.org/10.5281/zenodo.6160667 |
|
persistent identifier |
https://treatment.plazi.org/id/154FD01D-4B48-FFF7-FF6E-9116BB80CF03 |
|
treatment provided by |
Plazi |
|
scientific name |
Rhyacodrilus longichaeta |
| status |
sp. nov. |
Rhyacodrilus longichaeta sp. n.
( Figs 14–16 View FIGURE 14 View FIGURE 15 View FIGURE 16 )
Holotype. USNM 1202070, mature worm with sperm in the spermathecae, stained in hematoxylin, dissected and slide-mounted in Canada balsam.
Paratypes. USNM 1202071-74, 4 mature worms (2 with sperm in the spermathecae): 2 sagittally sectioned, 2 dissected and slide-mounted (23 June 2011); MNCN 16.03/3082: 1 whole mounted, unmated worm (15 May 1998), 1 dissected worm (6 April 2008). All from type locality.
Type locality. Guadalupe Creek, above Guadalupe reservoir, N37.1822° W121.8725°, Santa Clara Co., California (23 June 2011).
Other material. 18 partially mature specimens from the type locality: 2 mature unmated individuals, with sperm on male funnels, one sectioned (23 June 2011) and one dissected (20 June 2011). Other individuals with well-developed reproductive organs, with visible sperm sacs but sperm not fully developed: 2 whole mounts (15 May 1998); 5 dissected, (2 from 19 March 1999, and 2 from 9 April 2011); 8 whole mounts (25 April 2006). Additional unmounted worms from the type locality, all partially mature with visible sperm sacs: 6 in alcohol (19 March 1999), 14 in alcohol (25 April 2006). Immature specimens tentatively attributed to R. longichaeta : 7 immature whole mounts from Cooper River, above Cooper Lake, N47.4357° W121.1911°, Kittitas Co., Washington (1 November 1990); 3 immature whole mounts from the Merced River in Yosemite Valley, N37.7407° W119.5666°, Mariposa Co., California (6 October 1994 and 16 September 1995).
Futher material studied (not included in the description). Type series of Edmondsonia montana Brinkhurst, 1965 : Holotype USNM 32652 (1 slide), Paratype USNM 32653 (9 slides), all from Dog Lake, Yakima Co. Washington. An additional specimen attributed to this species, USNM 32980, from Sugar Island, Michigan.
Etymology. The specific name, longichaeta , refers to the extraordinarily long hair chaetae in segment II.
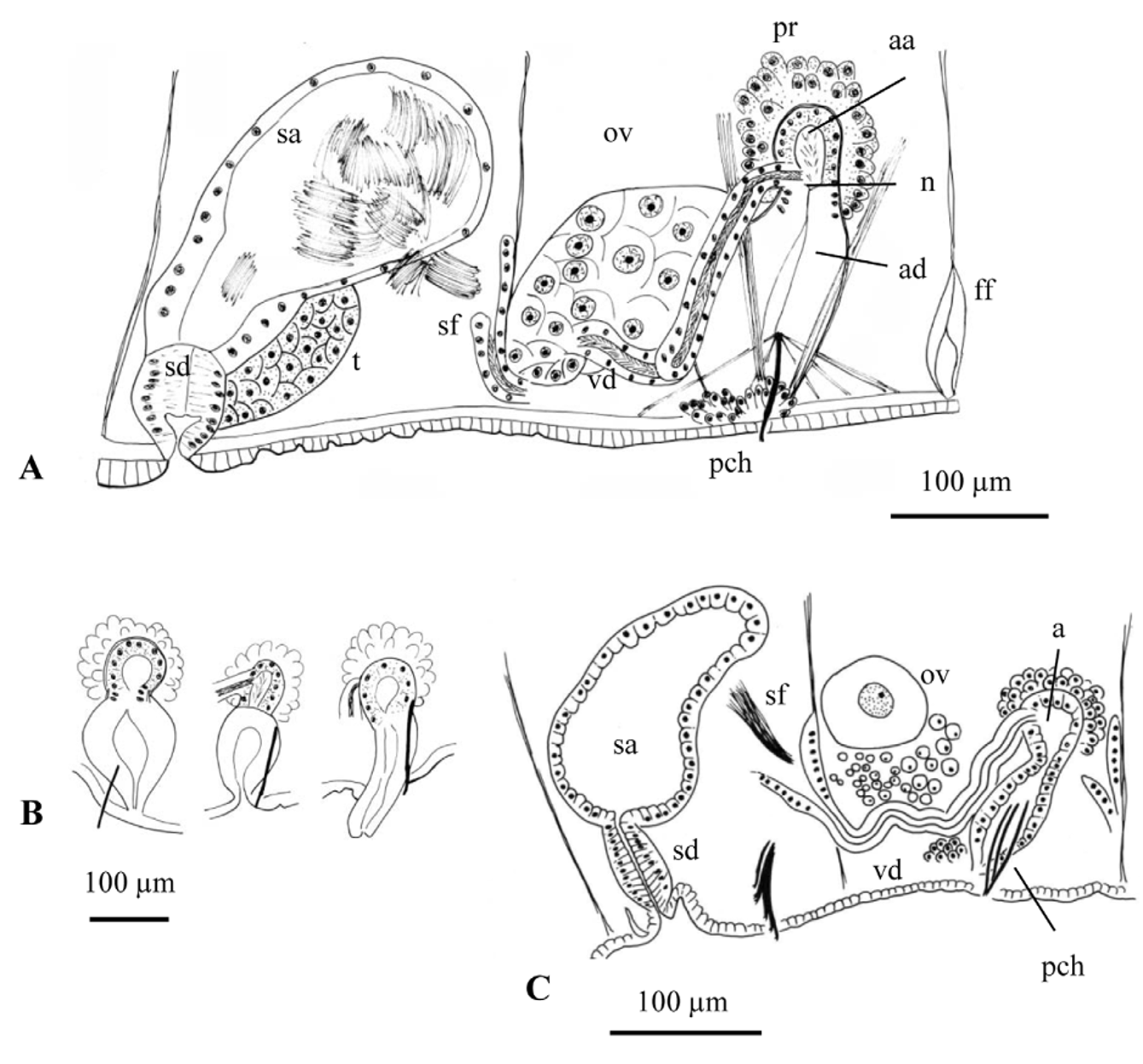
Description (from type locality specimens). Number of segments 48–95. Prostomium rounded to conical, 180–360 µm long ( Fig. 14 View FIGURE 14 A). Body diameter 0.42–0.74 mm in VIII. Secondary annulation usually absent in anterior segments, or a weak anterior ring in about IX–XI; variable in posterior segments, with 0–3 narrow rings. Mid-dorsal pores in intersegments (3/4) 4/5 to 6/7 (7/8). One pair spermathecal pores in the most anterior part of segment X, in line with ventral chaetae; two crescent-shaped grooves demarcate a median raised area between the pores, interpreted as a copulatory anchorage bridge. One pair male pores in the line of ventral chaetae in segment XI, usually opening on small round papillae; modified penial chaetae located closer to the body midline ( Fig. 14 View FIGURE 14 C). One pair female pores at intersegment 11/12, between lateral line and line of ventral chaetae.
Faintly hispid hair chaetae 1–4 per bundle in anterior segments (170–540 µm long), in segment II 1.4 to 2 times (620–960 µm) longer than the longest hairs in other preclitellar segments ( Figs 14 View FIGURE 14 A, 15F); hair chaetae present in most postclitellar segments, 0–1 per bundle, length less than 200 µm. Lateral hairs of hispid chaetae difficult to see in specimens from Guadalupe Creek (1000x usually required), but clearly visible at 400x magnification in Cooper and Merced River populations ( Fig. 15 View FIGURE 15 P). In anterior segments, 3–5 pectinate chaetae per bundle (96–144 µm long), with 4–7 intermediate teeth shorter than lateral teeth; 2–3 per bundle in posterior segments, commonly with very fine pectinations visible only at 1000x ( Fig. 15 View FIGURE 15 E–H). Upper teeth of anterior pectinates are slightly longer and thinner than, or equal to lower, although this varies even within the same bundle ( Fig. 15 View FIGURE 15 E); differences in size between lateral teeth are not as marked as in other populations (e.g. Cooper and Merced Rivers), or as in the type series of R. montana ( Fig. 15 View FIGURE 15 O, 16A, D, E). Ventral chaetae bifid; in anterior bundles 4–6(8) (96–149 µm long), with distal tooth thick and 1.5–2.5 times longer than proximal (longest in the most lateral chaetae within each bundle); 3–5 ventral chaetae in posterior segments (110–115 µm long) with distal tooth as long as but thinner than proximal ( Fig. 15 View FIGURE 15 I–M). In mature worms with sperm on the male funnels, ventral chaetae in segment X reduced in number (2–3 per bundle) and slightly larger (150–185 µm long), 1.2–1.8 times longer than chaetae in IX; the shape is also slightly modified, with distal tooth up to 3 times longer than lower ( Fig. 14 View FIGURE 14 B, 15N). At the male pore, (2)3(4) penial chaetae arranged more or less fanwise, simple-pointed or bifid ( Fig. 15 View FIGURE 15 D), 150–176 µm long, and associated with 2 strong dorso-ventral muscular strands ( Fig. 14 View FIGURE 14 E).
Pharynx with a well-developed dorsal muscular pad. Chloragogen cells start in segment VI and form a layer up to 27 µm high. Large coelomocytes (diameter 21–39 µm), nucleated and granulated ( Fig. 15 View FIGURE 15 A). Epidermis 11–18 µm high, circular muscle layer 5–8 µm thick and longitudinal muscles 10–30 µm thick. Clitellum 45–54 µm high dorsally, usually from about 1/ 2 X –XIII, may extend anteriad as far as the line of chaetae in IX. Pharyngeal glands from IV to VI, VII or VIII, in one individual back to the anterior part of IX.
Sperm funnel on septum 10/11. Vas deferens ciliated (ca. 390–750 µm long, 30–50 µm wide), narrowing (18– 30 µm) before subapical junction with atrium. Atrium club-shaped, total length 250–350 µm. Atrial ampulla elongate (length 110–168 µm, maximum diameter 54–74 µm), covered by a diffuse layer of prostate cells (up to 28 µm high), with thin musculature (ca. 3 µm thick). Atrial duct (96–192 µm long, 30–53 µm wide) not separated from ampulla by any constriction, although clearly distinguished by the absence of prostate cells, high columnar epithelium and prominent muscle strands that externally attach it to the ventral region around the male pores ( Fig. 14 View FIGURE 14 C, D, F). Sperm sac back to segments XVI–XXVII, egg sac back to XVI–XXVII.
Spermatheca with a short duct (55–108 µm long, 62–73 µm wide) that opens at the beginning of segment X, in the line of ventral chaetae. Spermathecal ampulla 168–305 µm long, of irregular sac-like shape, with unordered sperm ( Fig. 14 View FIGURE 14 C).
Anomalies. One individual from Guadalupe Creek had the genitalia shifted 3 segments forward (male pores in VIII) suggesting regeneration after fragmentation; one other specimen appeared to have a regenerating prostomium.
Distribution and habitat. Within Guadalupe Creek, R. longichaeta was found only in gravel-cobble sediments in riffle habitats above the confluence with Rincon Creek. Rhyacodrilus alcyoneus sp. n. was found only in riffles below this confluence, and neither has been found in downstream Guadalupe Creek sites, below the major reservoir. A benthic macroinvertebrate survey of Santa Clara Valley streams (Carter & Fend 2000) did not produce any other records of these species.
Remarks. Rhyacodrilus longichaeta sp. n. belongs to the group of Rhyacodrilus species with both hairs and pectinates, and also with several penial chaetae arranged fanwise and modified spermathecal chaetae. Within this group, R. svetlovi Sokolskaya, 1976 and R. alcyoneus sp. n. have a similar elongate, tubular atrium, with the atrial ampulla not separated from the duct by a constriction. The vas deferens of R. svetlovi is shorter than the atrium, and joins it apically. Differences from R. alcyoneus (described in the present paper) include the smaller size, the chaetal characteristics, the absence of ventral chaetal tubercula in IX and X, and the presence of hair chaetae in dorsal bundles of postclitellar segments. Both species have similar male ducts, with elongate-tubular atria and a weakly differentiated atrial duct; differences are not very pronounced, with the atrial duct less histologically differentiated in R. alcyoneus , and the simple male pore on a round papilla in R. longichaeta but opening within a fold in R. alcyoneus . Other Rhyacodrilus species with modified penial chaetae (but without spermathecal chaetae) and clubshaped to tubular atria are R. vasalatus and R. korjakovi , described by Semernoy (2004) from Lake Baikal. Of these two species, the male duct in R. longichaeta resembles that of R. vasalatus in having a vas deferens which narrows considerably before the atrial junction, and also in the high columnar epithelium of the atrial duct. However, R. vasalatus has only bifid chaetae, as does R. korjakovi when it is mature.
The lateral position of the female pores is uncommon in oligochaetes, where they typically have a more ventral position, in line with the ventral chaetae. Interestingly, this character is shared with other western American Rhyacodrilus species described here, i.e. R. saelonae , R. clio and R. alcyoneus .
Rhyacodrilus species with long hairs in II have generally been attributed to Rhyacodrilus (= Edmondsonia ) montana (Brinkhurst, 1965) , although elongate hairs in II were not mentioned in the original description. Furthermore, although diagnostic characters in Rhyacodrilus are largely based on details of the reproductive system, the original description of R. montana was based on immature worms, and only included characteristics such as the number and shape of chaetae and the abundance of coelomocytes (see Fig. 16 View FIGURE 16 ). The type series was collected at three sites in western North America: two from the northern Cascades Range in Washington (Dog Lake and Lake Tipsoo) and a distant California site (a pool at the edge of the Kings River); the holotype and a large series of paratypes were from Dog Lake. The species was transferred from the invalidated genus Edmondsonia Brinkhurst, 1965 to Rhyacodrilus by Brinkhurst and Cook (1966), based on the study of several populations of the Great Lakes region (eastern North America), but in that publication there is no comparison of chaetal characteristics with the type series and only limited information on the male duct.
Our examination of the R. montana holotype (USNM 32652) and all paratypes (USNM 32653) from the type locality has confirmed that the worms are immature. Hairs are not elongate in segment II of the holotype (485 µm in II and 345–530 µm in the following preclitellar segments) and of most paratypes. However, they are slightly longer in 3 paratypes (600–725 µm in II, and 500–650 µm in the following preclitellar segments, ratio of 1.1 between the longest in II and the longest in following segments); additionally, hairs are smooth, although they are originally described as "feathered hairs" (Brinkhurst 1965: 155). The anterior pectinates in R. montana have the distal tooth longer than the proximal (2 times longer in segments II–VIII of the holotype, about equal posteriorly), and ventral chaetae are pectinate ( Fig. 16 View FIGURE 16 B). The poor state of conservation of the tissues does not permit observations on internal anatomy, and even the chaetae are sometimes difficult to see. Accordingly, we propose that R. montana be considered species inquirenda due to its doubtful identity.
The long hairs in segment II are an obvious feature for species identification, since they can be clearly seen under the stereoscopic microscope. These chaetae, and the fanwise arrangement of the penial chaetae, were first reported for worms attributed to R. montana from the Great Lakes region by Brinkhurst and Cook (1966: 23) and used as diagnostic characters in subsequent literature (e.g. Brinkhurst & Jamieson 1971, Brinkhurst 1986, Kathman & Brinkhurst 1998). In a collection from this region (USNM 32980), reproductive organs are not visible (although one specimen has penial chaetae), and worms have very long hair chaetae in II (up to 1200 µm long, 1.8–2 times longer than the following ones), in contrast to the R. montana type series.
Anterior pectinates of R. longichaeta have only slightly longer distal teeth, in contrast to the distinctly longer and thicker distal tooth in the type material of R. montana ( Fig. 16 View FIGURE 16 A, D–E). Nevertheless, this character is usually difficult to interpret due to orientation of the pectinate chaetae, which are variably rotated. Immature rhyacodrilines with long hairs in segment II studied in other western localities (see other material above) are now provisionally ascribed to R. longichaeta . Anterior pectinates in these populations are variable; in some specimens the distal tooth is clearly longer than the lower, but not in others. Hispid hair chaetae are seen in some populations at 400x, but in individuals from Guadalupe Creek 1000x magnification is required, even with Nomarski microscopy. Hispid hair chaetae are rare in the genus, although they have been also described in R. hiemalis Ohtaka, 1995 .
Rhyacodrilus sodalis (Eisen, 1879) ( Figs 17 View FIGURE 17 , 18 View FIGURE 18 )
Ilyodrilus sodalis Eisen, 1879: 11 . Eisen 1885: 887, Pl. VI, Fig. 5 View FIGURE 5 .
Rhyacodrilus sodalis (Eisen) : Brinkhurst 1963: 53, 59.
Non Rhyacodrilus sodalis (Eisen) : Brinkhurst 1965: 144, Fig. 8 View FIGURE 8 E–K; Ohtaka 1995: Fig. 8 View FIGURE 8 B, C).
Neotype. USNM 1202075, dissected worm stained in Harris' hematoxylin and slide-mounted in Canada balsam.
Paraneotypes. USNM 1202076-77, 1 dissected worm stained in Harris' hematoxylin, 1 sagittally sectioned and stained with hematoxylin and eosin.
Further material, not included in description. Former neotype (USNM 32644) and 4 paraneotypes (USNM 32645 No.0004, 4 slides) of R. sodalis sensu Brinkhurst (1965) from Lake Tahoe, invalidated (see below).
Locality. Tributary to Mountain Lake, Camino del Mar, N37.7879° W122.4698°, San Francisco, California, USA, Eisen's original type locality of the species (31 May 2011).
Description. Number of segments 58–62. Body diameter about 0.55 mm at VIII, to 0.7 mm at clitellum.
Prostomium rounded. Epithelium in anterior segments up to 18 µm high. Clitellum annular from about 1/ 2 X to XII, absent only around male pores, up to 52 µm high, formed by small glandular cells ( Fig. 17 View FIGURE 17 A). Male pores in posterior part of segment XI, within a shallow, lenticular depression; spermathecal pores at the very beginning of segment X, close to intersegment 9/10; both male and spermathecal pores about in line with ventral chaetae. One pair female pores, lateral to the line of ventral chaetae. Anchorage bridge inconspicuous or absent, at most a small ridge of slightly thickened epidermis between spermathecal pores.
Dorsal bundles with hair and pectinate chaetae: 2–4 hairs in anterior bundles (the longest ones 190–410 µm), 1–2 thin hairs back to segment XVI–XXV, hairs absent posteriorly; 4–5 pectinate chaetae (83–132 µm) per bundle, with divergent lateral teeth of equal length and 4–5 intermediate teeth in anterior segments; 2–3 sigmoid pectinates in postclitellar segments (79–87 µm) with 3–4 short intermediate teeth, distal teeth shorter than proximal ( Fig. 18 View FIGURE 18 D, E), nodulus at 1/3 from the tip. Ventral bundles with 4–6(7) bifid chaetae in anterior segments (94–132 µm), 2– 5 in postclitelar segments (81–90 µm); teeth equally long or with distal tooth shorter and thinner than proximal; in most ventral bundles some chaetae have short thin intermediate teeth, the most posterior segments with only 1 bifid chaeta per bundle ( Fig. 18 View FIGURE 18 F–H). Median to the male pore a bundle of 3–4 penial chaetae (90–120 µm long, the longest in the middle of the bundle), bifid with short teeth, nodulus at 1/3 from the tip, arranged fanwise ( Fig. 18 View FIGURE 18 I, J).
Dorsal pharyngeal pad well-developed. Pharyngeal glands from segment IV to anterior part of segment VII. Numerous granulated and nucleated coelomocytes in body cavity (diameter 12–24 µm) ( Fig. 18 View FIGURE 18 A). One pair testes in segment X and one pair ovaries in segment XI. Sperm sacs extend forward into IX and back to segment XVIII– XX. Ovisac back to XIX–XXII. Paired ventral glands observed in segments V to X, best developed in segment IX, located behind ventral chaetae ( Fig. 18 View FIGURE 18 B).
One pair spermathecae, composed of a short bulbous duct (72–85 µm long, 52–72 µm maximum diameter) with columnar epithelium, and an irregular ampulla filled with bundles of sperm, characteristically folded (as in the original description of the species) around a strong muscular strand attached to the duct and to the dorsal body wall ( Fig. 17 View FIGURE 17 B: ms; 17C). One pair male ducts in segment XI. Sperm funnel on septum 10/11, followed by a long (to 700 μm), convoluted vas deferens, diameter 23–38 µm (thinner at both ends), apically joining the atrial ampulla ( Fig. 17 View FIGURE 17 B). Atrium with long duct (95–150 µm long, 27–30 µm wide), associated with several muscular bundles attached to the ventral body wall, and an elongated, curved ampulla (ca. 145–270 µm long, 70–125 µm maximum diameter). The ampulla may appear globular if compressed in whole mounted specimens, or elongated and curved (originally described as "crescent shape" [Eisen 1885: 888]) if relaxed or after removing from the segment through dissection ( Fig. 17 View FIGURE 17 B, D). As reported by Eisen (1885: 888), the atrium is "extremely pellucid" (transparent): atrial musculature very thin (ca. 1–3 µm), atrial epithelium thick (15–40 µm high), granular, and faint, although distinct in histological sections, with small, peripheral nuclei ( Fig. 17 View FIGURE 17 E). Ampulla covered by layer of prostate cells, 50–70 µm high, forming numerous clusters, joining atrium individually. In histological sections, a very short, narrow penis (?) was observed within the lumen of the atrial duct ( Fig. 17 View FIGURE 17 F). Anterior ends of spermatozoa in male duct and spermathecal ampulla have a distinctly wavy appearance (corkscrew shaped?) ( Fig. 18 View FIGURE 18 C).
Remarks. The species was described by Eisen (1879, 1885) as Ilyodrilus sodalis , from a small tributary to Mountain Lake in San Francisco (California). The species was transferred to the genus Rhyacodrilus by Brinkhurst (1963: 53, 59) based on atrial morphology, although regarded as a species dubia. Eisen's types collection disappeared after the earthquake in 1906 and a neotype was designated and described by Brinkhurst (1965). A sampling by Brinkhurst at the type locality was not successful in getting more material of the species (R.O. Brinkhurst, pers. comm. 2011), and the neotype was instead selected from Lake Tahoe. The new type series was however of limited value since neither male ducts nor spermathecae were observable (see Brinkhurst 1965: 145). Therefore, although the taxonomic status of the species has been considered uncertain (Brinkhurst 1986), this has not been an impediment to including the species in several guides, using keys based on chaetal characteristics. As a result, R. sodalis has been widely reported in North America (Kathman & Brinkhurst 1998).
Examination of Brinkhurst's neotype (USNM 32644) and paraneotypes from Lake Tahoe (USNM 32645 No.0004, 4 slides) did not provide any information on the structure of reproductive organs, probably due to the mounting media (most likely lactophenol). In the neotype, anterior pectinates have very long, thin teeth ( Fig. 18 View FIGURE 18 K, L) and there are 2 single-pointed penial chaetae, arranged in parallel. Lake Tahoe is a large, deep (499 m depth) water body, located at 1,897 m a.s.l. with highly oligotrophic waters (Gardner et al. 2000), while Eisen's original locality is in a very small, low elevation coastal seep, currently filled with vascular plants and draining an urbanized watershed. The linear distance between these two sites is about 230 km. The International Code of Zoological Nomenclature (ICZN 1999) states in Art. 75.3.6. that the paper in which a neotype is designated must state that "the neotype came as nearly as practicable from the original type locality and, where relevant, from the same geological horizon or host species as the original name-bearing type ". The poor condition of the Tahoe specimens, the lack of definition of the reproductive organs, and the fact that Brinkhurst's neotype series came from a water body very different in ecological and geological characteristics from the original indicates that the Lake Tahoe neotypes are invalid. Therefore we designate a new R. sodalis neotype from Eisen's type locality, adding details of morphological characters currently used in rhyacodriline descriptions. This description validates most of the morphological features formerly described by Eisen from the same locality.
The original description of R. sodalis gives little information on the shape of chaetae; however, the striated inner surface of the so-called "pseudo-comb-like spines" can be interpreted as the presence of intermediate teeth in dorsal chaetae, probably not clearly visible through Eisen's microcopy. Dorsal crochets are V-shaped with equal teeth in the San Francisco population (both in Eisen's description [1885: Fig. 5 View FIGURE 5 e,f] and in the present paper), instead of the long, thin teeth seen in the Lake Tahoe population. Large and abundant coelomocytes, a character that is diagnostic of the subfamily Rhyacodrilinae (Hrabĕ 1963b) , were not originally described by Eisen; they are present in the new neotype series, but this was likely not considered to be a diagnostic character at the time of the original description. There is a general agreement between Eisen's and the present descriptions of the spermatheca and male duct: the spermathecae have a short, bulbous duct, similar to many other Rhyacodrilus species, and an ampulla which appears twisted or folded; in the new material the ampulla is folded around a strong, dorso-ventral muscular strand originating at the spermathecal duct ( Fig. 17 View FIGURE 17 B, C). The male duct has a long, coiled vas deferens that apically joins the elongated atrium, and the long atrial duct opens in a simple pore. The conical structure that surrounds the atrial duct in the original description (incorrectly named oviduct by Eisen) is interpreted here as the covering of the atrial duct by numerous muscle strands attached to the ventral side of the body. The "pellucid" appearance of copulatory organs reported by Eisen is here interpreted as a vague or almost transparent atrial epithelium and very thin musculature. The most problematic issue is the lack of mention of modified penial chaetae in the original description of the species, a rather obvious character that is present in the specimens recently found in the type locality.
Based on the shape of the chaetae, presence of multiple penial chaetae but not spermathecal chaetae, the long vas deferens, and the atrial duct shorter than the ampulla, R. sodalis is most closely related to R. coccineus and a complex of similar species (see the key in this paper), some of which have been put in synonymy with the latter (e.g. Timm 1997). Rhyacodrilus sodalis has also been considered a junior synonym of other Rhyacodrilus species in this group. The synonymy of R. sodalis with R. sinicus Chen (e.g. Brinkhurst 1965, Brinkhurst & Jamieson 1971, Qi & Erséus 1985) is difficult to accept. In the original description by Chen (1940), the atrium is ovoid and the duct opens in a tegumentary fold, suggesting a closer relationship to R. coccineus . Rhyacodrilus sodalis has also been regarded as a possible junior synomym of R. coccineus (e.g. Timm 1997). Any proposal for synonymy of a species with the widespread R. coccineus requires a previous reconsideration of the taxonomic status of the latter, since at present the morphology of the species shows too much variability among the numerous descriptions by different authors in many parts of the world. At this point, we distinguish R. sodalis from R. coccineus mainly by the relatively long vas deferens joining the atrium apically, and by the more elongate, curved atrial ampulla described by Eisen and confirmed in the new material.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |