
Rhagasostoma rowei ( Brydone, 1906 )
|
publication ID |
https://doi.org/10.5852/ejt.2018.490 |
|
publication LSID |
lsid:zoobank.org:pub:BE023137-CC5E-4DC5-94F6-B549BB140361 |
|
DOI |
https://doi.org/10.5281/zenodo.3849968 |
|
persistent identifier |
https://treatment.plazi.org/id/03D587D7-965E-FF9F-F31C-F911FB61FA2C |
|
treatment provided by |
Valdenar |
|
scientific name |
Rhagasostoma rowei ( Brydone, 1906 ) |
| status |
|
Rhagasostoma rowei ( Brydone, 1906)
Figs 16–18 View Fig View Fig View Fig , Table 10 View Table 10
? Eschara ampullacea von Hagenow, 1839: 264.
? Eschara volgensis Eichwald, 1865: 193. Syn. nov.
Eschara rowei Brydone, 1906: 296 , fig. 6.
Eschara delarueana –? Marsson 1887: 69.
Eschara rowei – Brydone 1906: 296, fig. 6. — Brydone 1913: 249, pl. 8, fig. 12.
Onychocella (Eschara) cf. delarueana View in CoL – Voigt 1925: pl. 1, fig. 15.
Onychocella rowei View in CoL – Brydone 1930: pl. 28, fig. 3. — Voigt 1930: 455, pl. 15, fig. 11. — Voigt 1959: 9. — Voigt 1967: 41, pl. 18, figs 2–3. — Favorskaya 1996: pl. 4, fig. 1. — Koromyslova 2014a: pl. 9, figs 1–7, text-figs 1–4.
Woodipora mimosa ( Brydone, 1930) – Voigt 1967: 50, pl. 18, fig. 4.
Woodipora rowei – Schubert 1986 pars: 39, pl. 3, figs 5–6, 8, pl. 5, figs 1–3, 6–8, pl. 8, fig. 1.
non Eschara delarueana d’Orbigny, 1851: 105 , pl. 602, fig. 6–8, pl. 673, fig. 8
non Woodipora rowei – Schubert 1986 pars: 39, pl. 3, fig. 7, pl. 5, figs 4–5, pl. 8, figs 2–3.
Material studied
Holotype
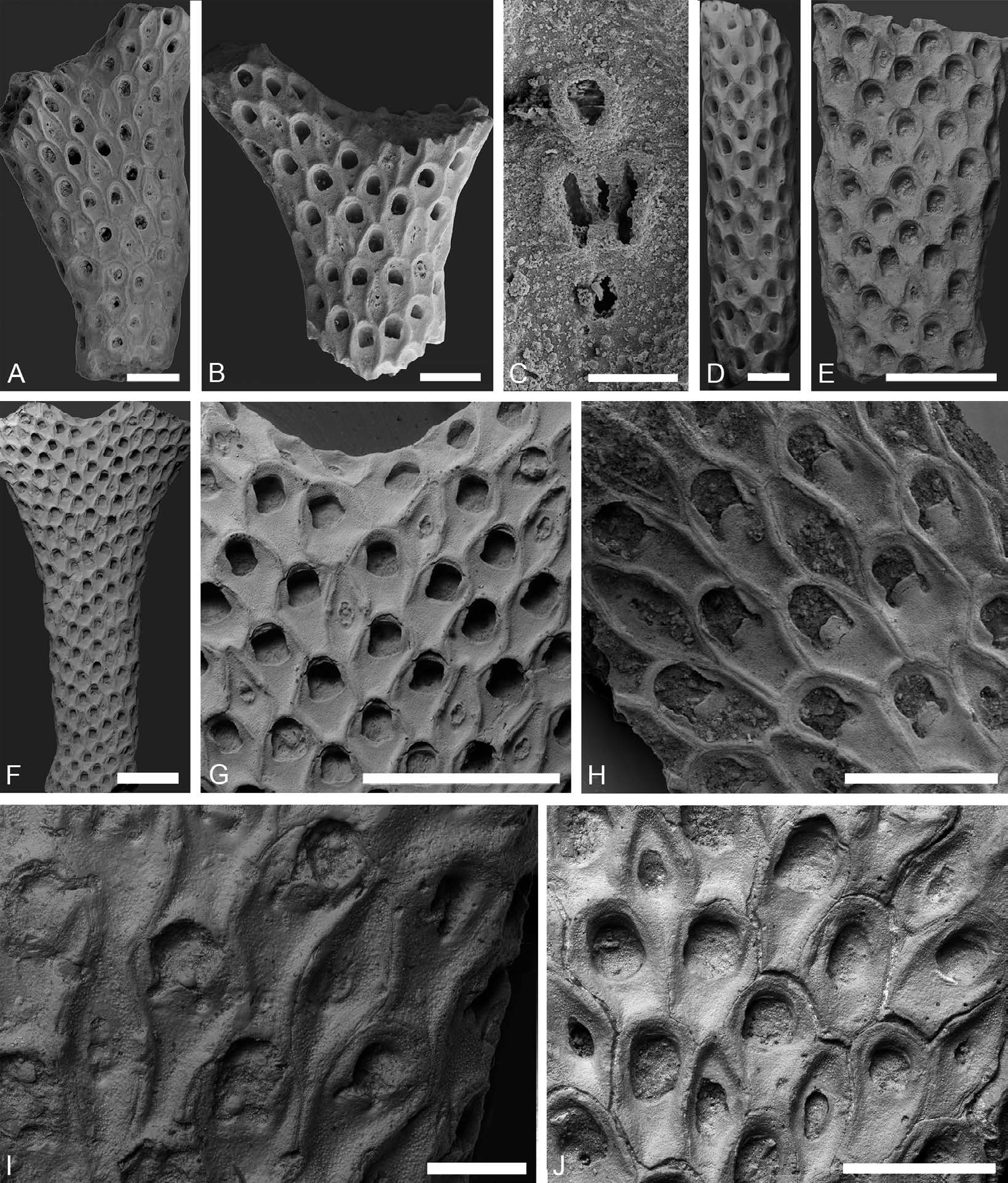
UNITED KINGDOM • England, Norfolk, Trimingham; early Maastrichtian ( Ostrea lunata Zone ); SM B36113 ( Fig. 16C View Fig ; not figured by Brydone 1906).
Additional figured material
RUSSIAN FEDERATION • 2 specs; Ulyanovsk Region; Late Cretaceous; PSM PSU 2/187 ( Fig. 16A View Fig ; figured by Eichwald 1868: pl. 8, fig. 9), MMI 15/49 ( Fig. 16B View Fig ; figured by Lahusen 1873: pl. 4, fig. 4).
UNITED KINGDOM • 2 specs; same data as for holotype; SM B36260 ( Fig. 16 View Fig D–E; figured by Brydone 1913: pl. 8, fig. 12), B36697 ( Fig. 16 View Fig F–G; labeled as O. mimosa in the Brydone Collection).
BELARUS • Grodno Region; erratic block of?late Campanian age in a quarry near Hrodna / Grodno (Гродна/ Гродно); PIN 2922 View Materials /204 ( Fig. 16 H View Fig ; figured by Koromyslova 2014a: pl. 9, fig. 6).
BELGIUM • Harmignies near Mons; late Campanian; SMF 29924 ( Fig. 17 View Fig A–B).
UZBEKISTAN • Southern Aral Sea Region; late Campanian, Cibicidoides voltzianus Zone ; TsNIGR Museum 26/12939 ( Fig. 17C View Fig ; figured by Favorskaya 1996: pl. 4, fig. 1).
TURKMENISTAN • Western Kopetdag; late Campanian; TsNIGR Museum 35/9757 ( Fig. 17 View Fig D–E; figured by Voigt 1967: pl. 18, fig. 3) • Tuarkyr; late Maastrichtian; TsNIGR Museum 36/9757 ( Fig. 18E View Fig ; figured byVoigt 1967: pl. 18, fig. 2) • Western Kopetdag; late Maastrichtian; TsNIGR Museum 32/9757 ( Fig. 18I View Fig ; figured by Voigt 1967: pl. 18, fig. 4).
KAZAKHSTAN • Emba River; late Campanian ( Belemnitella lanceolata Zone ); PIN 5502/3051 ( Fig. 17 View Fig F–G) • 2 specs; Mangyshlak Peninsula; Maastrichtian; PIN 3421/1009 ( Fig. 18 View Fig F–G), 3421/1008 ( Fig. 18H View Fig ) • Northern Aral Sea Region; Maastrichtian; PIN 5502/3050 ( Fig. 18J View Fig ).
GERMANY • 2 specs; Rügen ; early Maastrichtian; SMF 29931 ( Fig. 17 View Fig H–I), 29932 ( Fig. 18 A View Fig ) • Hamburg-Hummelsbüttel ; former brick factory; late Maastrichtian; SMF 29933 ( Fig. 18 D View Fig ) .
DENMARK • Island of Møn ; early Maastrichtian; SMF 29925 ( Fig. 18 View Fig B–C).
Other material
BELARUS • 15 specs; Grodno Region; erratic block of?late Campanian age in a quarry near Hrodna / Grodno (Гродна/ Гродно); PIN 2922/200 (figured by Koromyslova 2014a: pl. 9, fig. 1), 2922/201 (figured by Koromyslova 2014a: text-fig. 1c), 2922/208 (figured by Koromyslova 2014a: pl. 9, fig. 7), 2922/210 (figured by Koromyslova 2014a: pl. 9, fig. 3, text-figs 1a, b, 2), 2922/223 (figured by Koromyslova 2014a: pl. 9, fig. 5, text-figs 1g, 3a), 2922/224 (figured by Koromyslova 2014a: pl. 9, fig. 4, text-fig. 4e), 2922/226 (figured by Koromyslova 2014a: text-fig. 4d), 2922/227, 2922/234 (figured by Koromyslova 2014a: text-fig. 1d), 2922/231, 2922/255 (figured by Koromyslova 2014a: text-fig. 4ac), 2922/256 (figured by Koromyslova 2014a: text-fig. 4h), 2922/257 (figured by Koromyslova 2014a: text-fig. 4f), 2922/268, 2922/279 (figured by Koromyslova 2014a: pl. 9, fig. 2).
KAZAHKSTAN • 4 specs; Emba River; late Campanian ( Belemnitella lanceolata Zone ); PIN 5502/3052 to 3055 • Mangyshlak Peninsula; Maastrichtian; PIN 3421/1006.
GERMANY • 5 specs; Rügen ; early Maastrichtian; SMF 29926 to 29930 About SMF • 4 specs; Tornesch ; late Maastrichtian; SMF 29935 to 29938 About SMF • Hamburg-Hummelsbüttel ; former brick factory; late Maastrichtian; SMF 29934 .
Description
Colony rigidly erect, bifoliate, multiserial, branches 2.0–10.0 mm wide. Ancestrula and early astogeny not observed. Colony formed by pyriform and ovate autozooids and vicarious avicularia. Autozooids subrectangular with rounded distal ends; zooidal boundaries raised. Pyriform autozooids with a narrow proximal end, widening distally, the widening usually starting from the proximal margin of the opesia. In ovate autozooids, the proximal end is partly overlapped by the avicularian rostrum and the autozooid begins to widen considerably below the proximal margin of the opesia, usually at the boundary with an avicularium. Gymnocyst lacking. Cryptocyst slightly granulated, slightly depressed centrally and occupying half or more of the frontal surface of the autozooid, peripheral caverns lacking. Opesia terminal or subterminal. Opesial rim elevated, formed by projecting cryptocyst, bell-shaped due to small lateral projections (occlusor lamina) and opesiular indentations, between which there is a short or long, tongue-like projection of cryptocyst ( Figs 16A, C, E View Fig , 17G View Fig , 18 View Fig H–I), often broken, in which case the opesiular indentations are poorly defined and the proximal edge of the opesia is almost straight. In the complete absence of lateral projections and tongue the opesiae are subcircular ( Figs 17 View Fig C–E, H, 18A–B, D–G). Distal walls with two septula ( Fig. 16H View Fig ); septula in lateral walls not observed. Ovicells immersed, ooecia vestigial formed by the distal zooid, vizor-like, with slightly granulated surface, not protruding above colony surface ( Figs 16G View Fig , 17C, E, G View Fig , 18G, I View Fig ). Avicularia vicarious, rhomboidal. Rostrum conical in outline and symmetrical, usually with straight lateral walls; indentations or projections present at the rostral base. Three types of avicularia differing mainly in the length of the rostrum and rostral apex which partly overlaps the proximal end of the distal autozooid: avicularia with long rostra and long trough-like apices ( 130–210 µm); avicularia with short rostra and short trough-like apices ( 0–130 µm); and avicularia with rostra without projecting apices. Proximal part short, narrowing downwards, with convex lateral sides. Entire frontal surface of avicularium occupied by a fragile, slightly granulated cryptocyst containing five openings: two small, subcircular openings, distally and proximally, in between three slit-like, parallel openings ( Figs 17I View Fig , 18G View Fig ). Cryptocyst between openings frequently destroyed causing them to coalesce into a single opening of variable outline. Intramural reparative budding of autozooids and closure plates not observed. Intramural reparative budding of avicularia may be present in some or all avicularia depending on the colony (see Koromyslova 2014a), they are observed within host avicularia, numbering as many as three intramural buds, all apparently having the same polarity as the host avicularium.
Kenozooids not observed. Both the inner and outer sides of branches of colonies show concentrations of autozooids in which the opesiae are subcircular and the proximal part is shortened or completely absent, and deformed avicularia that are irregularly ovate to rhomboidal in shape and possess an elongated opening at the centre.
Remarks
Eschara volgensis Eichwald, 1865 from the Late Cretaceous of Simbirsk (now Ulyanovsk, Russian Federation) was regarded as possibly conspecific with Rhagasostoma rowei by Voigt (1967). Voigt, who examined the Eichwald Collection housed at the Palaeontological and Stratigraphical Museum at the Faculty of Geology, St Petersburg State University, found that the original material of this species from Simbirsk, Ulyanovsk Oblast, Russia differs from R. rowei only in the opesia being smaller and having a slightly visible tongue-like projection of the cryptocyst. Unfortunately, the specimen of E. volgensis (PSM PSU 2/187 and PSM PSU 2/188) could not be found during our visit to the collections. However, a photograph of the specimen made by Prof. E. Voigt in 1963 is reproduced here ( Fig. 16A View Fig ). This species was redescribed by I.I. Lahusen (1873) whose collection is stored in the Museum of the Mining Institute, St Petersburg. The specimen of E. volgensis (MMI 15/49) could not be examined using SEM, but a microphotograph is shown instead ( Fig. 16B View Fig ). The Lahusen material of E. volgensis is from the vicinity of Yazykov village in Simbirsk Province, Ulyanovsk Oblast. He stated that the sample described by Eichwald (1865) was not completely identical with his samples. Our findings show that the sample described by Lahusen is conspecific with Rhagasostoma rowei .
In a monograph on the development of different species of Woodipora Jullien, 1888 from the Coniacian to the Maastrichtian, Schubert (1986) transferred Brydone’s species rowei to the genus Woodipora . He did not regard the presence of opesiules or opesiular indentations as being diagnostic for species and concluded that rowei and mimosa were synonymous, differing simply in the preservation of the opesiules (for a more detailed summary regarding a possible synonymy of rowei and mimosa , see the remarks for R. mimosa ).
Rhagasostoma rowei has bell-shaped autozooidal opesia because of the small lateral projections and opesiular indentations separated by a short projecting tongue of cryptocyst. Opesia with a straight or rounded proximal edges are formed when the lateral projections and cryptocystal tongue become broken-off. When the cryptocyst located between the opesiules in autozooids of R. mimosa is destroyed, the opesia takes a shape similar to that of R. rowei . Thus, this similarity is related to the state of preservation and it is evident that pristine colonies of R. rowei have opesiular indentations whereas those of R. mimosa have opesiules.
We have restudied the type material from the collections of R.M. Brydone and also material from the collections of I.I. Lahusen, E. Voigt and T.A. Favorskaya. The specimens labelled as O. mimosa (SM B36697) from the Brydone Collection and (TsNIGR Museum 32/9757) from the Voigt Collection belong to R. rowei because the autozooids lack opesiules. Usually R. rowei is characterized by colonies that widen distally with numerous avicularia. The large number of avicularia is due to the fact that avicularia occur at the start of the new zooidal rows needed to widen the branches. From these avicularia, ovate autozooids are budded distally, from which in turn pyriform autozooids are budded ( Koromyslova 2014a). However, there is one specimen of R. rowei from the Maastrichtian of Turkmenistan ( Fig. 18E View Fig ; figured by Voigt 1967: pl. 18, fig. 2) that differs from the holotype and many other specimens in having almost parallel-sided branches and rare avicularia. Additionally, some large colonies are almost parallel-sided at the base with rare avicularia but widen distally where numerous avicularia are present ( Figs 17 View Fig A–B, 18F–G). It can be assumed that the samples described by Voigt (1967) and other similar specimens ( Fig. 18D View Fig ) were broken-off from the bases of such colonies and hence have few avicularia. Ovate autozooids, which are budded from avicularia, are also rare and often poorly expressed. Obviously, R. rowei was usually characterized by stem-like colonies, which are almost parallel-sided at the base and have few avicularia, but become fan-like distally where numerous avicularia are present. Furthermore, colonies of this species can occasionally have bifurcating branches ( Figs 16 View Fig B–C, F, 18A–B, F). Entire colonies are rare and usually we can see only a small part of colonies, which were broken-off. Schubert (1986) mentioned the presence of R. rowei in his distribution list from Royan in southwest France. However, it would be rather surprising to find this species in the Aquitaine Basin, as the bryofauna described so far from the Aquitaine Basin differs considerably from the northern Chalk Sea bryofauna. In his synonymy list and in his remarks, Schubert (1986) placed, following Voigt (1930), Marsson’s (1887) Eschara Delarueana (non d’Orbigny, 1851) and Voigt’s (1925) Onychocella cf. delarueana in synonymy with R. rowei . However, this material is derived from the early Maastrichtian of Rügen ( Marsson 1887) and the late Maastrichtian of the ‘Blue Hills’ (Blaue Berge) in eastern Germany ( Voigt 1925). The true Rhagasostoma delarueana (d’Orbigny, 1851) from Royan, however, is not conspecific with R. rowei . Since no samples of R. rowei were found among the material studied by Schubert (1986) labelled “Royan”, this locality has to be deleted from the distribution list of R. rowei .
Schubert (1986) erroneously considered all material from Møns Klint in eastern Denmark as late Maastrichtian. However, no sediments from the late Maastrichtian crop out at Møns Klint (cf. Mutterlose et al. 1998) and the samples studied by Schubert (1986) are in fact labelled as early Maastrichtian.
A specimen comparable to R. rowei has recently been described as ‘ Onychocella ’ sp. in Koromyslova et al. (2018a) from the early Maastrichtian of the Aktolagay Plateau, western Kazakhstan. The Aktoalagay species, however, is encrusting and only very poorly preserved.
Distribution
Late Santonian France: Vasterival near Sainte-Marguerite-sur-Mer, Normandy ( Schubert 1986). Late Campanian Belgium: Harmignies near Mons. Belarus: Quarry near Hrodna / Grodno (Гродна/ Гродно), Grodno Region ( Koromyslova 2014a). Turkmenistan: “Kredin” gorge in the Kopet Dag Mountains NW of Aşgabat ( Voigt 1967). Uzbekistan: southern Aral Sea Region, Republic of Karakalpakstan ( Favorskaya 1996). Kazakhstan: Emba River. Early Maastrichtian Denmark: Møns Klint, Sjaelland ( Schubert 1986). Germany:Hemmoor and Lüneburg, Lower Saxony ( Voigt1967);Saturn quarry near Kronsmoor, Schleswig-Holstein ( Schubert 1986); Island of Rügen, Mecklenburg-Vorpommern (von Hagenow 1839; Voigt 1930). Kazakhstan:Along the Chobda river, and Kenderlyshor, Mangystau Region ( Voigt 1967). United Kingdom: Trimingham, Norfolk, England ( Brydone 1906). Late Maastrichtian Denmark: Kongsted (Faxe Kommune; Schubert 1986). France: Chef-du-Pont, Normandy ( Schubert 1986). Germany: Blaue Berge near Dessau-Rosslau, Saxony-Anhalt ( Voigt 1925), Tornesch, Hamburg-Hummelsbüttel. Turkmenistan: Koimat gorge and a gorge W of the “Kamyschli” gorge in the Kopet Dag Mountains NW of Aşgabat, Tuarkyr ( Voigt 1967). Maastrichtian Kazakhstan: northern Aral Sea Region, Mangyshlak Peninsula.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Microporoidea |
|
Family |
|
|
Genus |
Rhagasostoma rowei ( Brydone, 1906 )
| Koromyslova, Anna V., Taylor, Paul D., Martha, Silviu O. & Riley, Matthew 2018 |
Woodipora mimosa ( Brydone, 1930 )
| Voigt E. 1967: 50 |
rowei
| Brydone R. M. 1906: 296 |
rowei
| Brydone R. M. 1913: 249 |
| Brydone R. M. 1906: 296 |
delarueana
| Marsson T. 1887: 69 |