
Chorilaena, Endl.
|
publication ID |
https://doi.org/10.1071/SB22018 |
|
DOI |
https://doi.org/10.5281/zenodo.10988422 |
|
persistent identifier |
https://treatment.plazi.org/id/865587A8-FFF5-FFAA-FFFA-8A936C38FAB1 |
|
treatment provided by |
Felipe |
|
scientific name |
Chorilaena |
| status |
|
Chorilaena View in CoL and Rhadinothamnus
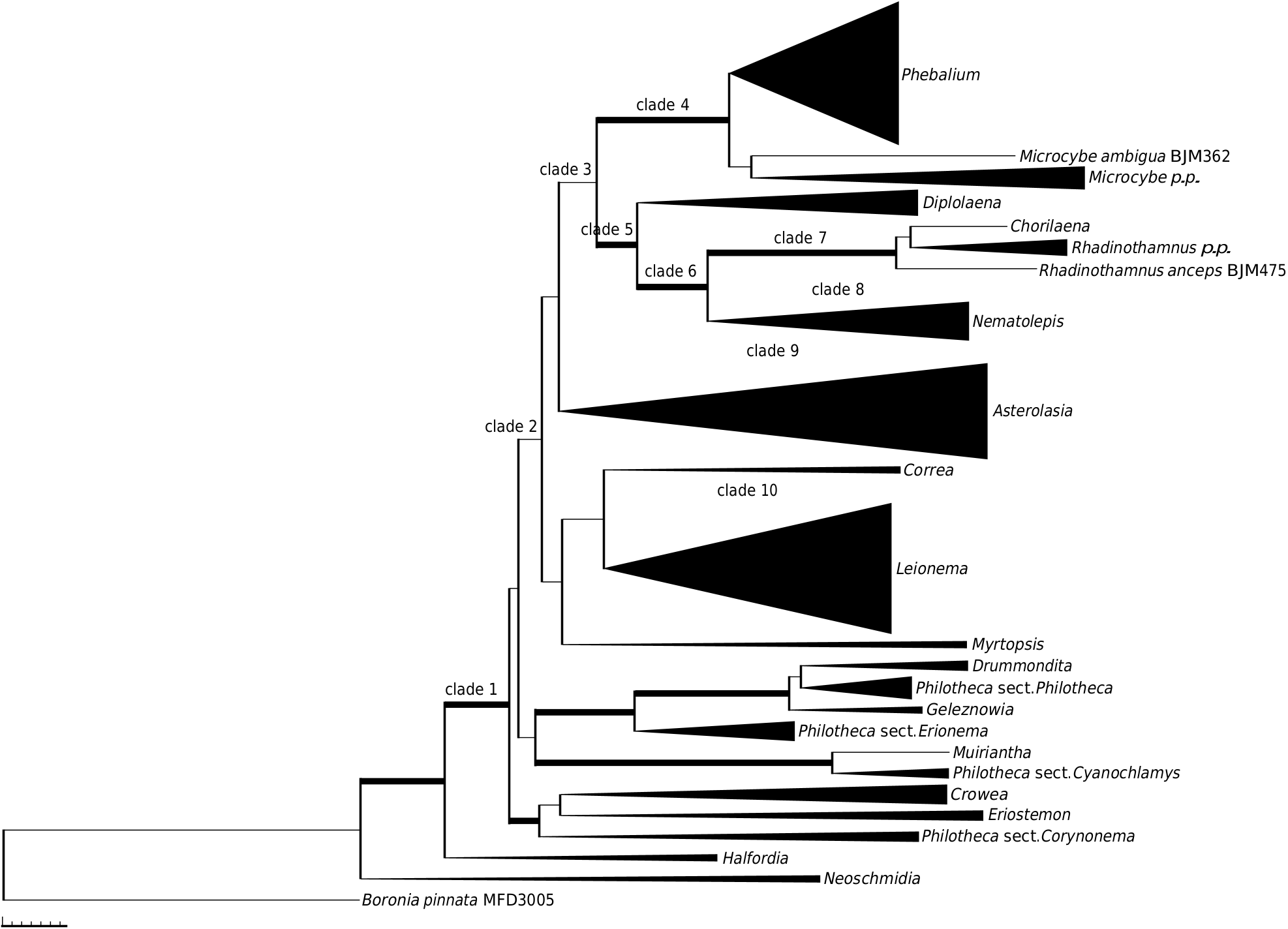
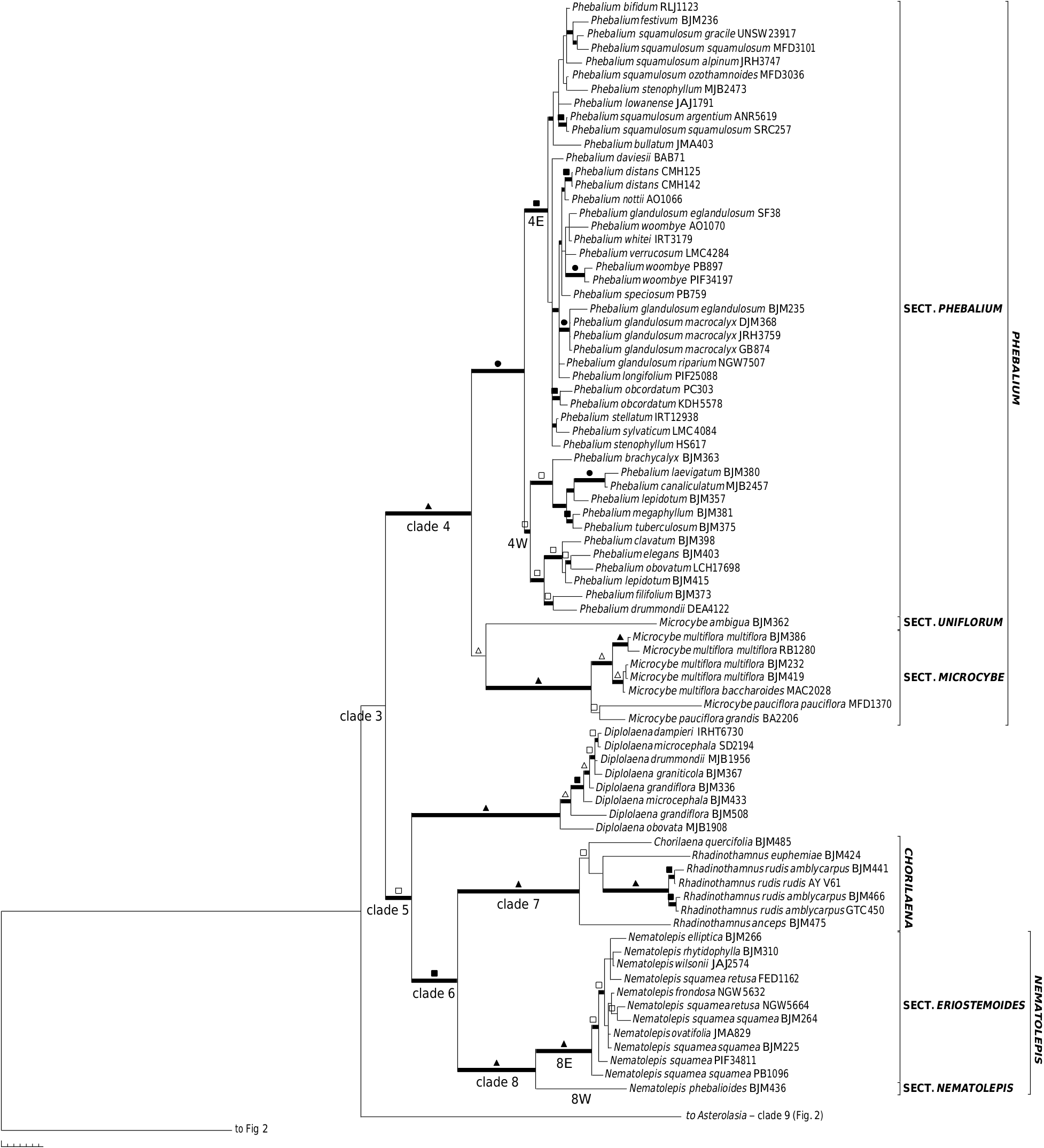
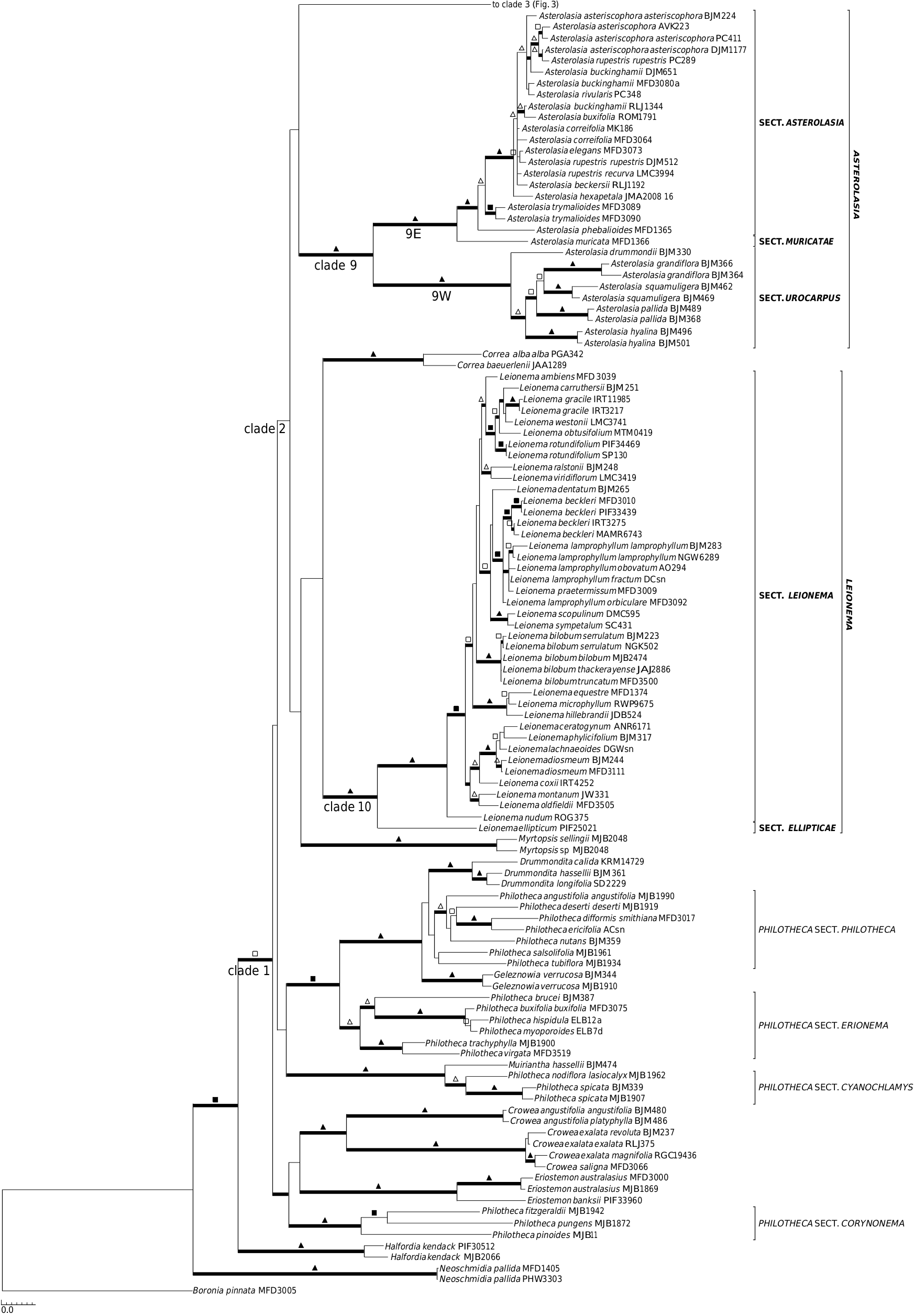
A close relationship between Chorilaena (monotypic, sampled) and Rhadinothamnus (all 3 species and 2 of the 3 subspecies of R. rudis sampled) has been postulated before, because the genera share a unique seed type that has two persistent cartilaginous strands on the adaxial surface between which the aril is attached, as well as the hemispherical calyx, valvate petals, and non-glandular anther apiculum ( Wilson 1970, 1971, 1998 b, 2013 f; Mole et al. 2004; Armstrong 2013). The results presented here ( Fig. 1 View Fig , 3 View Fig , S 1 View Fig , S 2 View Fig ) indicate that retaining the two genera, as currently circumscribed, is not well justified. Mole et al. (2004), referring to this issue, indicated that broadening the concept of Chorilaena to include all species of Rhadinothamnus would create an extremely morphologically diverse genus and their preferred option was to retain Rhadinothamnus and Chorilaena as monotypic and raise the former Phebalium section Gonioclados (containing R. anceps and R. rudis ) to generic rank, despite the section not having good support in their analysis. The necessary nomenclatural changes were not made. In the analyses presented here, there is no support for this proposition and no strong support for any species–pair relationships within this clade. The length of the branches in this clade are comparable to that found in other genera. The four species being retained in one genus, an expanded Chorilaena , would create a morphologically diverse genus, in habit, leaf form, and in inflorescence and flower morphology. The inflorescence and flower diversity are, presumably, driven by differences in pollination systems. The morphological diversity seen in the Chorilaena + Rhadinothmanus clade is no more diverse than that seen in other Australasian genera of Rutaceae where various pollination systems appear to have evolved (see discussion below on pollination).
Results presented here and by Mole et al. (2004) indicated that there are the following four taxonomic choices for these four species: (1) acknowledge the close relationship of the four species and expand Chorilaena to include all four; (2) because there are no clear relationships identified among the four species, have four monotypic genera, which would require two novel genera to be formally described; (3) raise Phebalium section Gonioclados to genus level, as proposed by Mole et al. (2004), for the superficially morphologically similar R. anceps and R. rudis , despite this relationship not being supported by molecular data; or 4, recognise that the exact relationships of C. quercifolia Endl. and R. anceps to the other species are not well supported at this stage and maintain the status quo until they are more clearly established. Here, we adopt Option 1 to reflect the data and make the appropriate nomenclatural changes below.
The close relationship of the expanded Chorilaena to Nematolepis is strongly supported (1.00 PP, 95% JK) in our analyses, confirming the results of Mole et al. (2004). Wilson (1970) had also considered Phebalum section Eriostemoides (= Nematolepis ) and P. section Gonioclados (= Chorilaena ) to be closely related.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Zanthoxyloideae |