
Pseudostomella mandela, Todaro, M. Antonio, Perissinotto, Renzo & Bownes, Sarah J., 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3905.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:3C2F37A5-262F-4C36-BF66-FD3254E2F147 |
|
DOI |
https://doi.org/10.5281/zenodo.6094136 |
|
persistent identifier |
https://treatment.plazi.org/id/C8FA4EBD-5939-4F79-AD7F-4CB02FE184CA |
|
taxon LSID |
lsid:zoobank.org:act:C8FA4EBD-5939-4F79-AD7F-4CB02FE184CA |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudostomella mandela |
| status |
sp. nov. |
Pseudostomella mandela View in CoL n. sp.
Figs. 6–9 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9
ZooBank lsid:zoobank.org:act:C8FA4EBD-5939-4F79-AD7F-4CB02FE184CA
Type locality. South Africa, KwaZulu-Natal, Cape Vidal, iSimangaliso Wetland Park (Latitude 28°07’18” S; Longitude 32°33’43” W); at about 1.5 m water depth, in fine to medium, well sorted sand (mean grain size, 0.246 mm; sorting, 0.46). Values of salinity and pH of coastal waters around the time of sampling were 36.8 and 8.4, respectively, with temperatures ranging from 27–28° C.
Other locations. Umhlanga (Durban) at 5 m water depth in fine, moderately well sorted siliceous sands (mean grain size, 0.228 mm sorting, 0.51); St. Lucia beach, at mid-tide level in medium-sized, moderately sorted, siliceous grains on a high-energy sandy beach (see Todaro et al. 2011b).
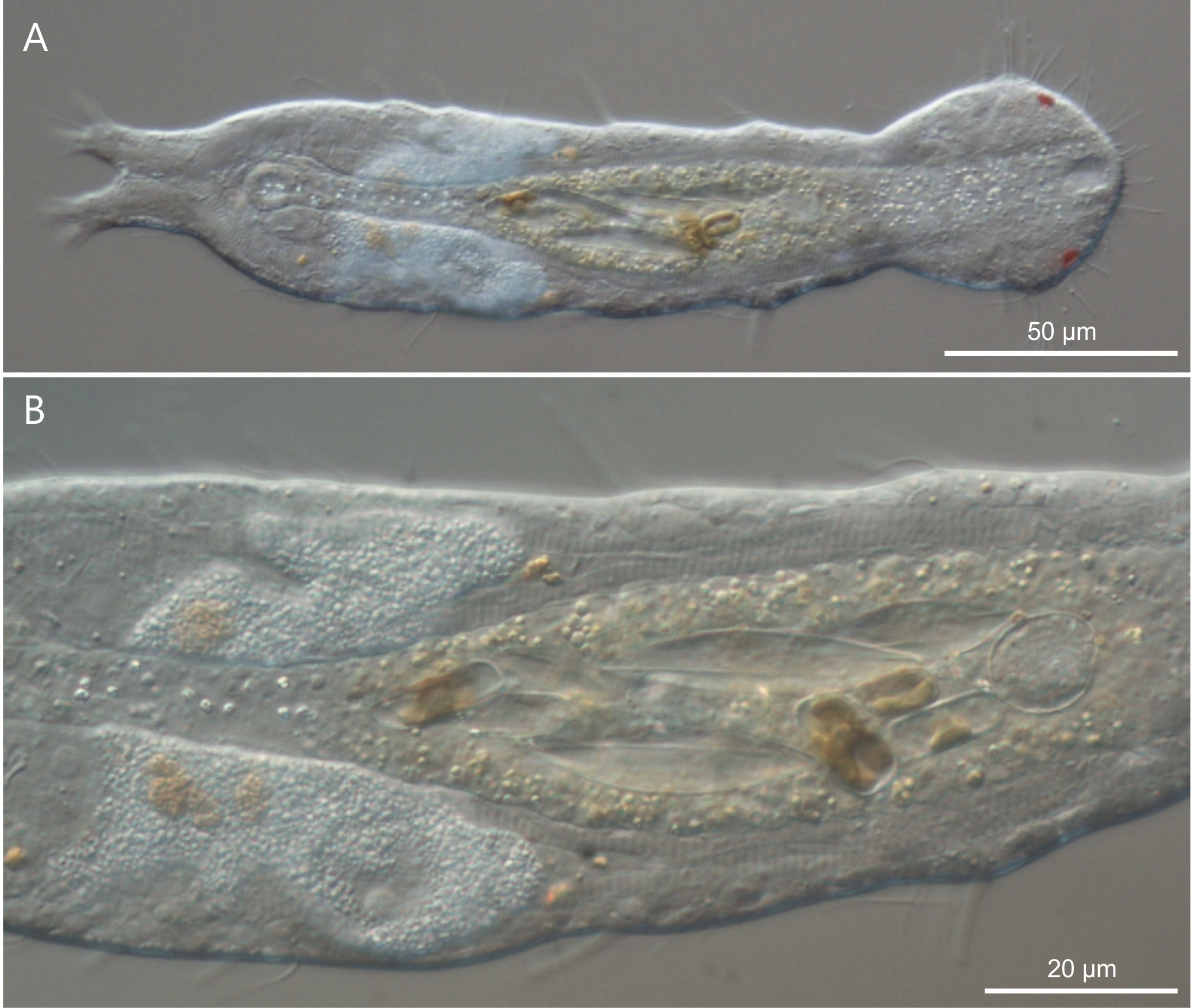
Type specimens. Holotype: the 477 Μm long adult specimen shown in Figure 7 View FIGURE 7 , no longer extant (International Code of Zoological Nomenclature, Articles 73.1.1 and 73.1.4), collected on 2 February 2013 (MAT legit).
Additional material examined. Five adult specimens, four collected by the author from the type locality and one from Umhlanga, all were observed alive and are no longer extant. Three further identified specimens were fixed in alcohol and are kept in the collection of the first author.
Ecology. Frequency of occurrence: common in sediment along the ocean shore of the iSimangaliso Wetland Park, usual in southern sites near Durban. Abundance: prevalent to numerous in sub-littoral sediment at 1.5 m, scarce in deeper and littoral sediments where found.
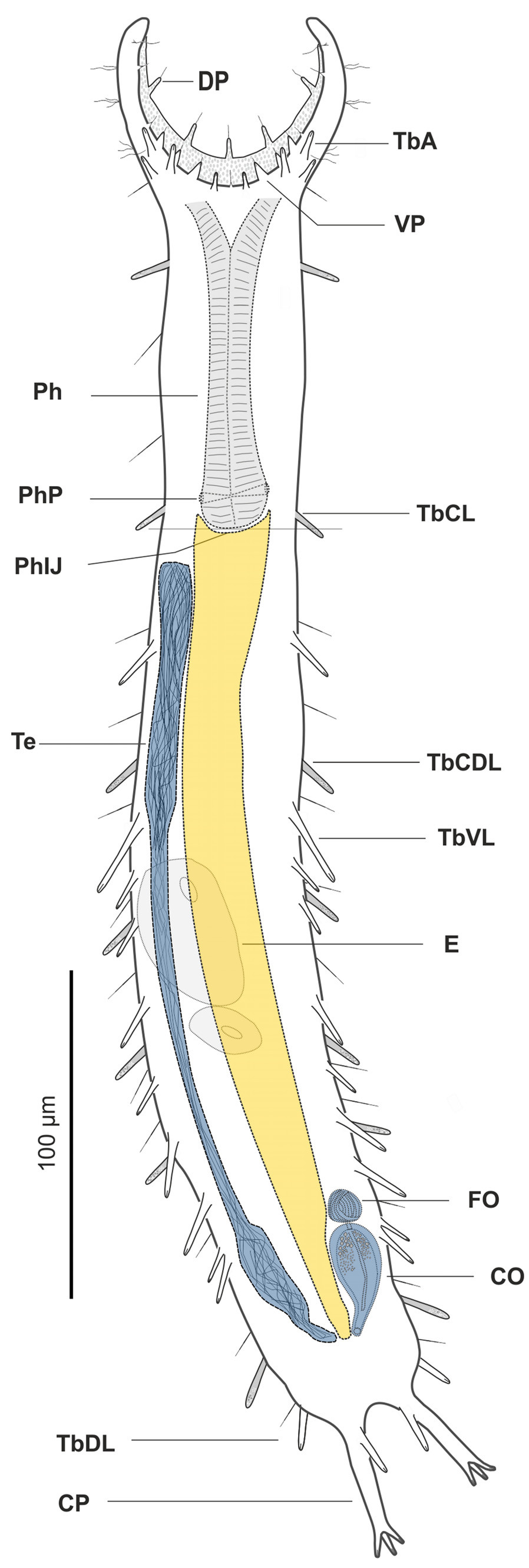
Diagnosis. A Pseudostomella with an adult TL up to 481 µm; pharynx length up to 105 µm, with pharyngeal pores at base. PhIJ at U33; body slender, with fine lines and elongate, furcate caudum. Head with mid-sized fleshy preoral palps curving forward; palps showing few sensory hairs and provided with 5 and 6 papillae on the dorsal and ventral border respectively. Sensory hairs sparse but evenly spaced on the body, forming two columns from about U11 to U89; epidermal glands barely visible, asymmetrically scattered along most of the length of the body. Cuticular armature of medium-size tetrancres on whole dorsal and ventrolateral surface. Adhesive tubes: TbA, 4 per side, a small one medially and 3 larger ones laterally, forming an arc; TbDL, 1 per side, inserting on lateral margin of the posterior trunk region at U90; TbVL, 11 per side, of considerably variable length, irregularly spaced along the intestinal region from U40 to U81; the last two tubes appearing very close to each other but not sharing a common base; TbP, 4 per side, 2 + 1 at the end of each caudal pedicle the other one flanking each pedicle medially; cirrata tubes, 7 per side, 2 smaller ones inserting laterally along the pharyngeal region at U16 and U32 respectively, and 5 larger ones inserting dorso-laterally on the trunk from U48 to U87; the last one particularly large. Ventral locomotory cilia: a continuous field of transverse rows covering sparsely the entire ventral surface except the anogenital area. Reproductive system: testis on the right body side, sperm duct noticeably swollen at the confluence of the ano-genital area; caudal organ inverted, pyriform, at U80; frontal organ, small, bladder-like, at U77; maturing eggs mid-dorsally above the intestine.
Etymology. The species is dedicated to the late Nelson Rolihlahla Mandela , the first democratically elected President of South Africa and 1993 Nobel Peace Prize awardee.
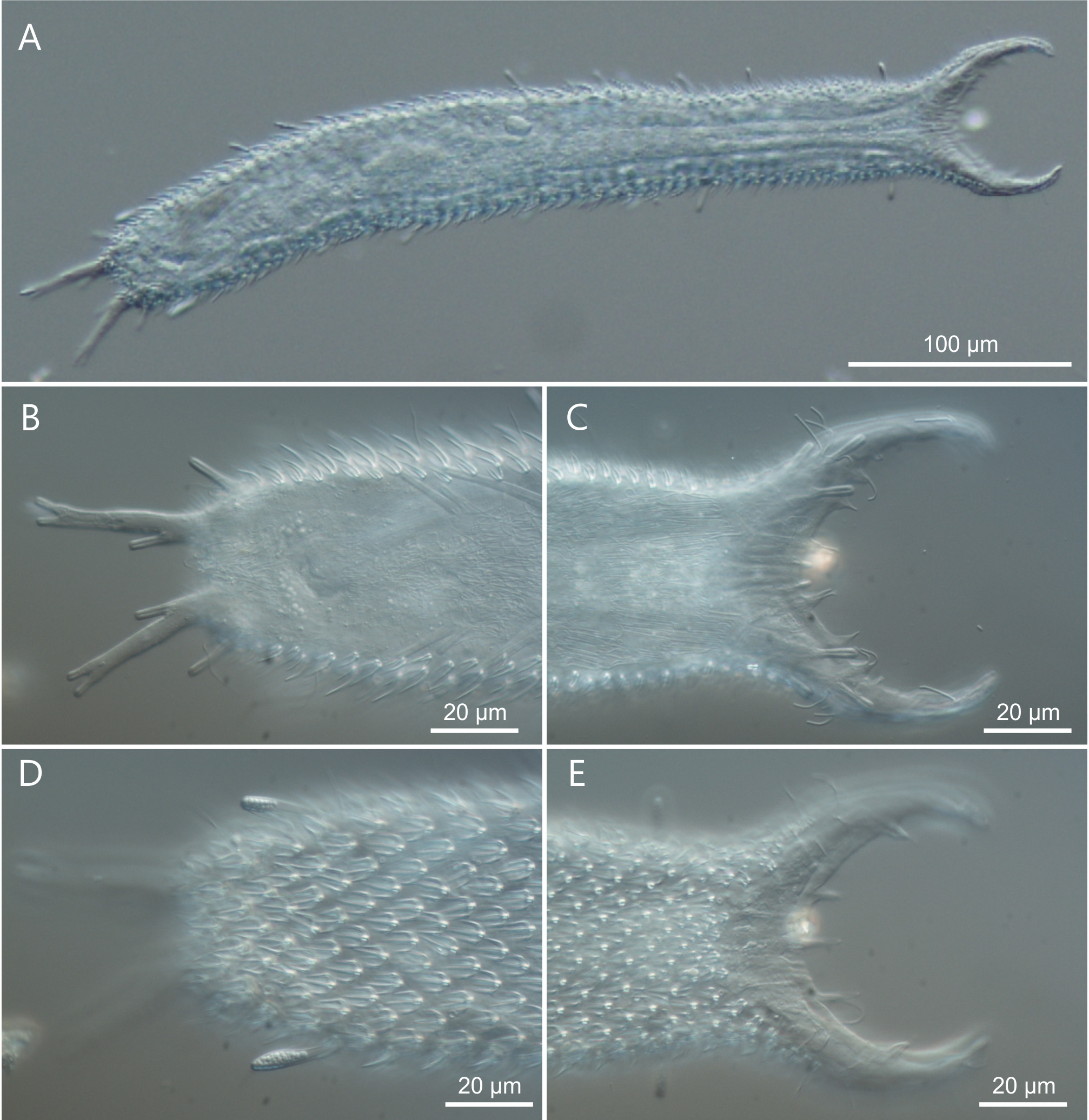
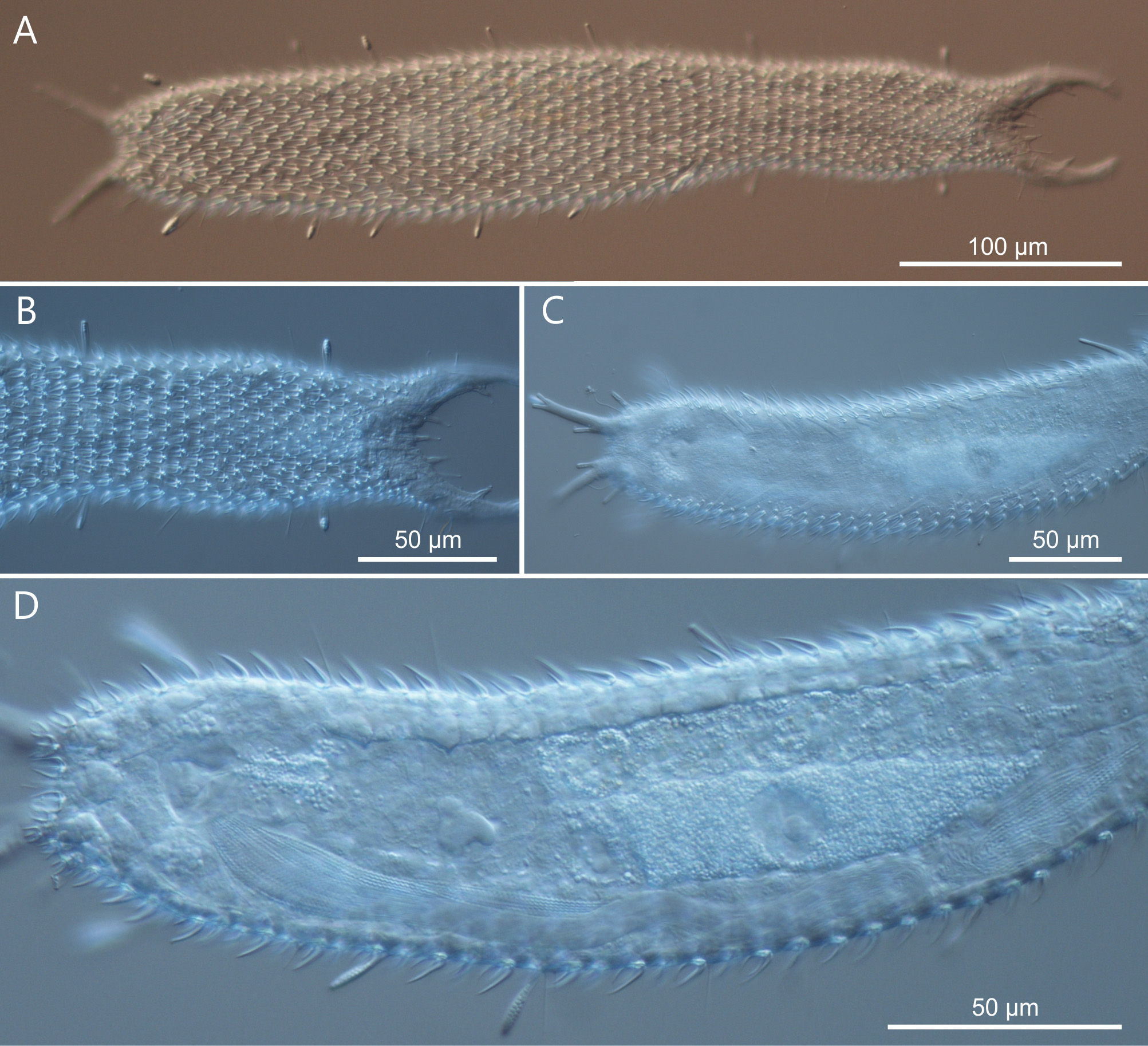
Description. The description is mainly based on the adult holotype specimen, 477 µm in total length, shown in Figure 6 View FIGURE 6 . Body somewhat slender, a little swollen in the posterior pharyngeal region and at the base of the 44 µm long caudal pedicles. Pharynx 104 µm in length, measured from the ventral border of the oral opening to the pharyngeo-intestinal junction; pharyngeal pores near the base at U31; pharyngeo-intestinal junction at U33; widths of neck\PhIJ\trunk\caudal base are 45\41\57\28 µm at U22\U33\U51\U91, respectively. Head with well developed, fleshy preoral palps, curving ventromedially; the dorsal border projecting just beyond the ventral. Sensory hairs and papillae present on dorsal and ventral borders of the preoral palps; hairs are scattered on the dorsal, lateral and ventral surface of the palps; dorsally there are five papillae, nearly of the same length (5–7 µm), symmetrically arranged along the inner border of the palps in a 2 + 1 + 2 pattern; ventrally, there are six clearly recognizable papillae, 3–6 µm in length, symmetrically arranged more centrally about the inner border of the palps in a 3 + 3 pattern; all papillae bearing one or two short sensory hairs at their tip; other sensory cilia form lateral columns that are more or less evenly spaced from U11 to U89; individual hairs are 12–15 µm in length. A variable number of additional papillae, much smaller in size (1–3µm), can be found ventrally along the inner border of the palps. Epidermal glands barely visible, variable in shape (oval to oblong) and size (5–8 µm in diameter), asymmetrically scattered along most of the length of the body.
Cuticular armature. Mid-sized tetrancres with delicate, nearly straight lines, taller than wide (3 x 2 µm – 10 x 7 µm) on the whole dorsal and ventrolateral surface; posteriorly ancres do not extend onto the caudum.
Adhesive tubes. TbA, 4 per side, a small one medially (3 µm in length) and 3 larger ones laterally (7–8 µm in length), forming an arc at U09-U10; TbDL, 1 per side (14–16 µm in length), inserting on the lateral margin of the posterior trunk region at U90; TbVL, 11 per side, of different size, irregularly spaced along the intestinal region from U40 to U81; 5 tubes are visibly larger (23–29 µm in length) and 6 are smaller (11–17 µm in length) than the others; the last two smaller tubes appear very close to each other yet they seem to originate independently, i.e. do not share a common base; TbP, 4 per side, 2 + 1 (5–6 µm in length) at the end of each pedicle of the furcated caudum and the other one (10–11 µm in length) flanking each pedicle medially.
Ventral locomotory cilia. A continuous field of transverse rows covering sparsely the entire surface except around the ano-genital area at U85.
Reproductive system. Testis on the right body side, sperm duct noticeably swollen at the confluence of the anogenital area; caudal organ inverted, pyriform (35 µm long x 15 µm wide), at U80; frontal organ, bladder-like (10–12 µm in diameter) at U77; maturing eggs mid-dorsally above the intestine.
Variability and remarks. The body length of 6 living specimens ranged from 465 to 481 µm (mean = 476.6 µm SD = 5.3 µm), all of them were mature (i.e. showed at least the testis filled with sperm). One specimen (TL = 480 µm) showed an asymmetry in the TbA by virtue of a supernumerary fifth tube on the right side ( Fig. 9 View FIGURE 9 ); the adhesive tubes of the TbVL series showed some variability in number, depending on individuals, ranging from10 to 13 tubes; however, in all specimens the last two tubes were very close to each other with the longest of the two being the terminal one. Although the last two tubes appear to originate independently under DIC optics, a final word on this matter would necessitate a SEM survey.
In a previous study, we reported on a juvenile specimen of Pseudostomella found at St Lucia beach (iSimangaliso Wetland Park, Todaro et al. 2011b). The cuticular covering, made up of tetrancres, the rather long caudal pedicles and the same area of the finding leave no doubt that the current adult specimens and the juvenile found previously belong to the same species. This is despite some differences regarding the presence of TbL in the juvenile but not in the adults, which bear cirrata tubes instead. Cirrata tubes have been described for several thaumastodermatid species e.g., Tetranchyroderma (Todaro 2002) and are generally thought to convey the secretions (probably with a repellent function) produced by the epidermal glands externally. The densely packet droplets clearly visible inside the cirrata tubes of P. mandela n. sp. (see Fig. 7 View FIGURE 7 D), resembling the epidermal droplets recently described for Ptychostomella lamelliphora Todaro, 2013 (see Todaro 2013, Fig. 4 View FIGURE 4 D), seem to support this hypothesis. The reported disparities between juveniles and adults highlight the significant ontogenetic morphological variations experienced by Pseudostomella specimens during their life, and once again caution researchers on using only mature specimens for a reliable identification and/or species description.
Taxonomic affinities. Within Thaumastodermatinae , the genera Tetranchyroderma and Pseudostomella , the number of prongs forming the peculiar scales called ancres, which composes the body cuticular armature, has been regarded as the single most useful taxonomic trait to classify species (e.g., Lee & Chang 2002; Todaro 2002; but see Todaro et al. 2011a). Consequently, in the genus Pseudostomella three basic species groups are envisaged based on the type of pronged spines i.e., species characterized by triancres (3 prongs), tetrancres (4 prongs) or pentancres (5 prongs). Based on the type of ancres, Pseudostomella mandela n. sp. approaches six other species all characterized by a tetrancrous covering: P. andamanica Rao, 1993 , P. indi ca Rao, 1970, P. koreana Lee & Chang, 2002 , P. longifurca Lee & Chang, 2002 , P. m al ay i ca Renaud-Mornant, 1967 and P. ro s c o vi t a Swedmark, 1956 (see Todaro 2012). However, based on the number of dorsal papillae (5) on the preoral palps, the new species is most similar to P. longifurca and P. indica . The number and distribution of the adhesive tubes is useful in discriminating the three taxa e.g., P. longifurca bears 5 TbA and 7 TbP per side and P. i n di c a 2 TbA and 5 TbP, in contrast with the new species that exhibits 4 TbA and 4 TbP per side. P. mandela n. sp. is further distinguished as the only species in the genus that possesses cirrata tubes (7 pairs).
The finding of an additional new taxon seems to support the idea that Pseudostomella species appear to have a relatively restricted geographic range, at least compared to the wide distribution and cosmopolitan nature of many other gastrotrichs (cf. Todaro et al. 1996; Artois et al. 2011; Curini-Galletti et al. 2012; Kånneby et al. 2012; Kieneke et al. 2012).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Thaumastodermatinae |
|
Genus |