
Promesacanthus eppleri, Hanke, 2008
|
publication ID |
https://doi.org/ 10.5281/zenodo.5378575 |
|
persistent identifier |
https://treatment.plazi.org/id/40637552-9818-A161-26F2-0A84EDD4408D |
|
treatment provided by |
Marcus |
|
scientific name |
Promesacanthus eppleri |
| status |
sp. nov. |
Promesacanthus eppleri n. sp. ( Figs 2-7 View FIG View FIG View FIG View FIG View FIG View FIG )
Mesacanthidae gen. et. sp. nov. – Wilson et al. 2000: 139.
Acanthodiformes , undescribed species – Hanke 2002: 1072.
“New mesacanthid” – Hanke 2002: 1079.
HOLOTYPE. — UALVP 41860 , a fairly intact, small specimen showing details from head to tail, preserved with left side exposed.
ETYMOLOGY. — Eppleri, honoring Allan Eppler, great friend, fellow scholar, and maritime naturalist.
MATERIAL EXAMINED. — UALVP 41672, 42651, 42652, 42653, 43027.
TYPE LOCALITY AND HORIZON. — All specimens are from the Early Devonian (Lochkovian) MOTH locality, GSC 69014, section 43 of Gabrielse et al. (1973); the fish bearing horizon is between 430-435 m in the section (as measured in 1996); in dark grey, argillaceous limestone.
DIAGNOSIS. — A small acanthodiform with laterally compressed body; Meckel’s cartilage ossified as single element; simple, single articulation between Meckel’s cartilage and palatoquadrate; head with enlarged, platelike, irregularly-shaped scales; ornamented sclerotic plates present; enlarged interorbital plates absent; blade-like, ornamented hyoidean gill covers present; one pair of short prepectoral spines present; fin spines ornamented with a thick longitudinal rib at the leading edge and a posterior field of fine, parallel longitudinal striations; scapulocoracoids with cylindrical scapular blade and triangular coracoid portion; procoracoids with round dorsal process articulating with similarly shaped fossa on anteromedial face of scapulocoracoids; dorsal fin spine base positioned ahead of anal fin spine base; thin rhombic endoskeletal basal plate posteroventral to dorsal fin spine; scales small with smooth flat crowns; largest body scales at base of dorsal fin spine.
DESCRIPTION
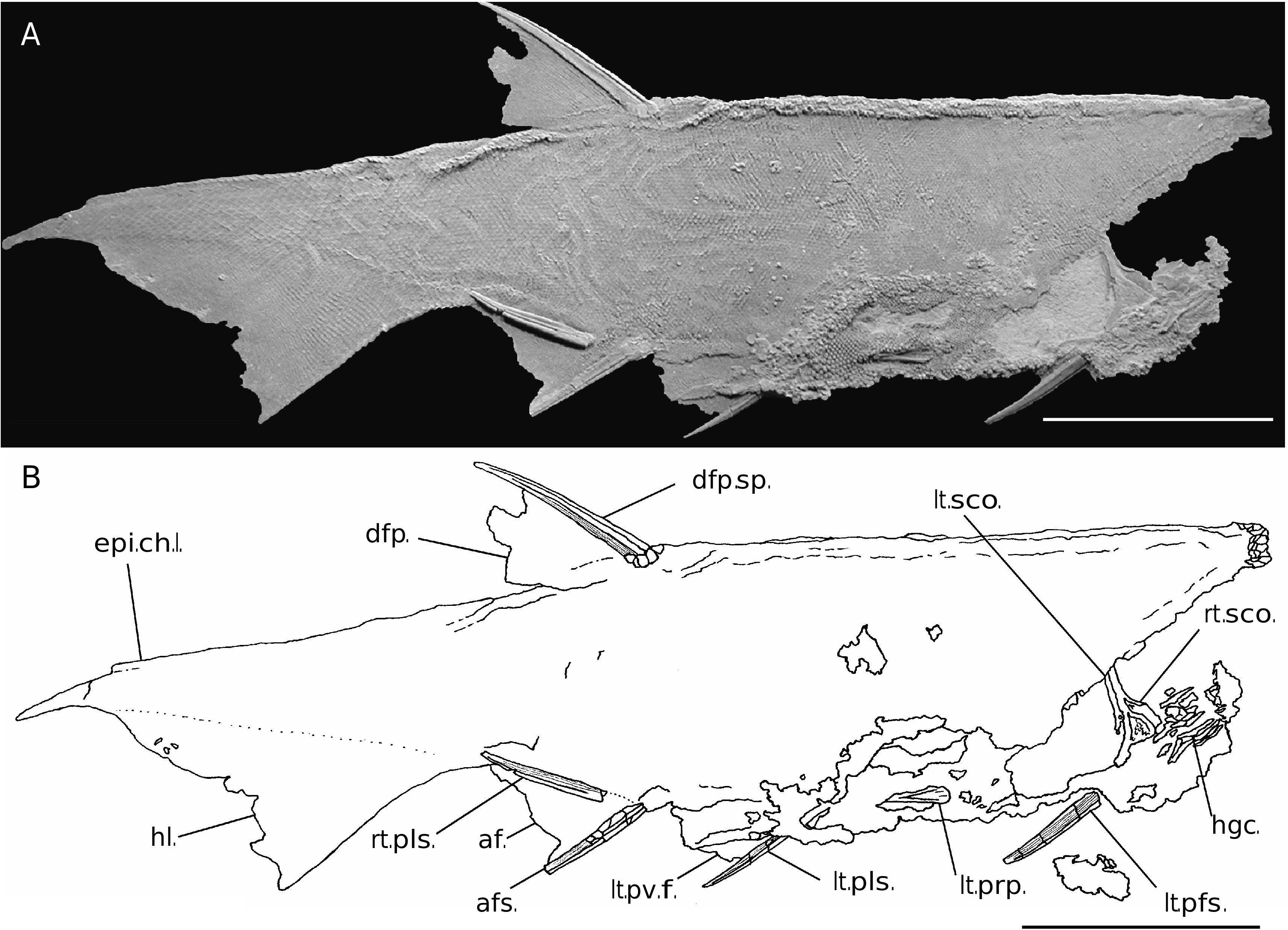
Most of the description that follows is based on specimens UALVP 43027 and 42652. Promesacanthus eppleri n. gen., n. sp. is a small, elongate acanthodian with a body depth/length ratio of approximately 0.17 based on the holotype (UALVP 41860), to 0.19 estimated by joining the larger specimens 42652 and 43027 at the pectoral girdle ( Figs 2-4 View FIG View FIG View FIG ). The body was laterally compressed in cross section, based on the fact that all known specimens are preserved on their side. Ridges are formed along the dorsal and ventral margins of the body fossils where scales along the back and belly collapsed together during decay and settling of the carcass. The body also is elongate, and tapers gradually to the caudal peduncle. The trace of the main lateral line extends along the body from an anterior position over the branchial chamber, posteriorly to the lower half of the caudal peduncle ( Fig. 2 View FIG ).
Head and jaws
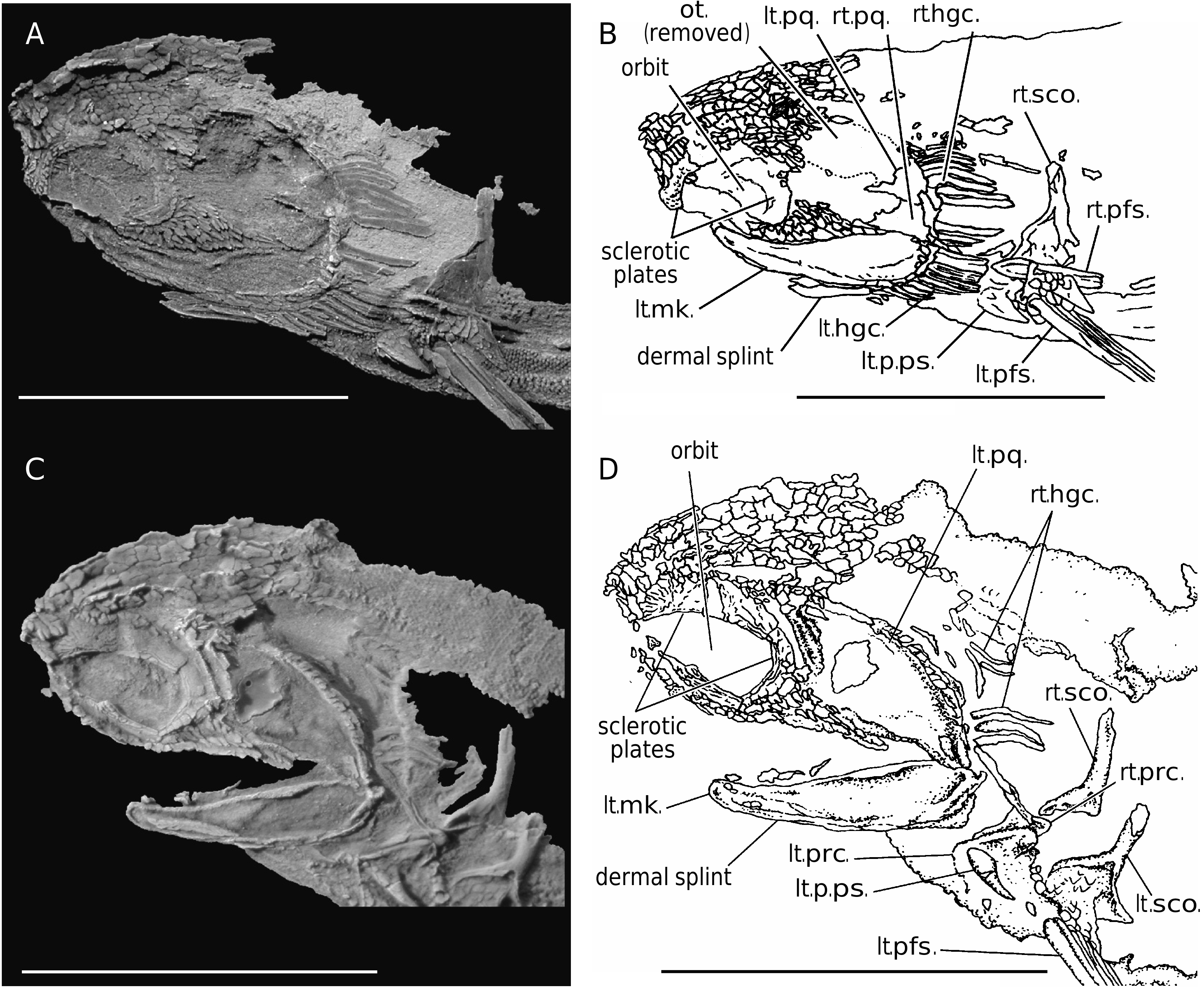
The head is broader than the body. The heads of most specimens are preserved as an oblique compression as the armored head settled during decay, whereas the body with less girth, preserved as a near perfect lateral compression ( Figs 3 View FIG ; 4 View FIG ). The braincase is unossified, although the position of the otic portion of the braincase is indicated by two patches of statoconia ( Figs 2 View FIG ; 4B View FIG ). The rostrum is short and overhangs the mouth, but specialized nasal scales and/or enlarged interorbital plates are absent. The head likely was widest across the otic portion of the braincase, and the large eyes must have been only slightly separated medially.
The orbits are large and surrounded by typical head scales along the anterodorsal margin ( Figs 4 View FIG ; 5 View FIG A-D). The anterior margin of the orbit is positioned anterior to the symphysis of the lower jaw, and the eyes contain four thin sclerotic plates; a gap in the sclerotic plates in the anteroventral quarter of the eye ( Fig. 4 View FIG ) may have contained a fifth plate. The rim of each sclerotic plate (nearest the pupil) is ornamented with fine, flat, round- to irregularly-shaped tubercles ( Fig. 5A, C View FIG ). The rest of the external surface of each plate is ornamented with broad flat ridges that radiate towards the back of the eye.
The rostrum and the dorsal surface of the head are covered with enlarged, square to irregularly-shaped scales ( Figs 4 View FIG ; 5A, B View FIG ). The crowns of the head scales have a smooth lobate ornamentation, and the basal surface is flat to slightly concave.
Asymmetrical scales are present posteroventral to the orbits, and these cover the autopalatine portion of the palatoquadrate ( Figs 4 View FIG ; 5C, D View FIG ). The crowns of these cheek scales have a narrow anterior end, an irregularly shaped, lobate posterior edge, and a flat to slightly concave basal surface. Each lobe on the trailing edge of the cheek scales ventral to the orbit, forms short, broad processes, whereas scales posterior to the orbit have elongate finger-like trailing processes. Larger cheek scales may have up to five trailing processes. Similar scales are found along the ventral edge of the Meckel’s cartilage, anterior to the hyoidean plates, and along the extrapalatoquadrate crest of the upper jaw ( Figs 4 View FIG ; 5C View FIG ).
Meckel’s cartilages each are preserved as a single unit in the larger specimens; the lower jaws of the smallest specimen (holotype) are not obvious. The lower jaw is slender anteriorly, and gradually deepens posteriorly, narrowing again at the jaw articulation ( Fig. 4 View FIG ). Meckel’s cartilages are supported by a dermal splint ( Figs 4 View FIG ; 5C View FIG ), and have a fairly wide articular cotylus, but lack a prominent preglenoid process ( Fig. 4C, D View FIG ). Both Meckel’s cartilages and the palatoquadrate cartilages are calcified, and have a fine-grained surface texture. None of the jaw elements have been thin-sectioned to determine the type of tissue present.
The palatoquadrate is formed from at least two elements, a larger quadrate portion and a smaller metapterygoid portion ( Fig. 4C, D View FIG ). The presence and structure of the autopalatine portion cannot be confirmed because of scale cover posteroventral to the orbit. The palatoquadrate is large, extending posterior to the otic portion of the braincase. A low extrapalatoquadrate crest is present, and it is covered with scales that are similar to those posteroventral to the orbit and along the ventral margin of Meckel’s cartilage ( Fig. 4C, D View FIG ). The metapterygoid portion of the palatoquadrate is sub-triangular and has an anterodorsal thickening presumably for articulation with the braincase. The foramen for the mandibular branch of the trigeminal nerve may have been present, but it is not visible due to damage on both specimens showing jaw structure; it is possible that the thin edges of the foramen flaked away during preparation of the specimen. The otic articular surface is covered by scales and cannot be described without additional specimen preparation. There is a broad, low flange just anterior to the jaw articulation that passes medial to the dorsal edge of Meckel’s cartilage. This flange may correspond to a prearticular process. The articular process is wide and forms a simple, single articulation surface for the upper and lower jaws. All specimens of P. eppleri n. gen., n. sp. lack teeth.
The gill arches are not mineralized. The extent of the branchial chamber is estimated from the space between the angle of the jaws and the position of the pectoral girdle. The branchial chamber is compact and the operculum is reinforced by at least eight thin, blade-like, ornamented hyoidean plates above the jaw articulation ( Fig. 4A, B View FIG ). These hyoidean plates probably covered most of the dorsal half of the branchial chamber. Approximately ten plates are present ventral to the jaw articulation, and presumably these reinforced the ventral portion of the branchial chamber to the isthmus ( Figs 4A, B View FIG ; 5F View FIG ). The hyoidean plates are smooth and unornamented on the visceral surface, and the external surface is ornamented with overlapping ridges forming a nested series of chevrons.
Pectoral girdle
The scapulocoracoid has a thin, straight, elongate scapular blade which is elliptical in cross section, and a broad triangular coracoid portion for articulation with the procoracoids and the pectoral fin spine ( Figs 3 View FIG ; 4 View FIG ). Each scapulocoracoid is ossified as a single unit. The coracoid portion of the scapulocoracoid has a convex anterior edge and concave posterior edge in lateral view, and the scapular blade is nearly vertical. Each procoracoid is positioned anteromedial to the coracoid portion of its respective scapulocoracoid ( Fig. 4 View FIG ). The dorsal process of the procoracoid is rounded and articulates with a similar shaped concavity on the anteromedial face of the coracoid portion of its respective scapulocoracoid. The ventral portion of the procoracoid is covered in all available specimens and cannot be described without additional preparation.
The pectoral fin spine of P. eppleri n. gen., n. sp. is slender, curves near the tip ( Figs 2 View FIG ; 3 View FIG ), and is the longest fin spine on the body. The pectoral spines are reinforced with a single rib along the leading edge and a fairly thick posterolateral rib per side. The posterior portion of the pectoral fin spine is ornamented with a field of five to six fine longitudinal striations that continue along the entire spine, parallel to the leading rib. Fin spine ribs and striations are smooth and lack nodular ornament. The pectoral fin spine is shallowly inserted into the body wall and has an elongate, narrow basal opening ( Fig. 4A, B View FIG ).
A single prepectoral spine is present just anterior to the base of each pectoral fin spine ( Figs 2 View FIG ; 4 View FIG ). The prepectoral spines are short, stout, lack ribs and have fine longitudinal striations. Promesacanthus eppleri n. gen., n. sp. is the first acanthodiform known to have prepectoral spines. Pinnal and lorical plates are absent.
The bases of the prepectoral and the pectoral fin spines are surrounded by scales which have large, flat to convex crowns ( Fig. 4A View FIG ). The pectoral fin web either had no scale cover, or it was formed from fine scales which were lost from all available specimens.
Dorsal fin and spine
Promesacanthus eppleri n. gen., n. sp. has one dorsal fin, and this likely corresponds to the posterior dorsal fin of non-acanthodiform acanthodians ( Figs 2 View FIG ; 3 View FIG ). The dorsal fin spine is second in length to the pectoral fin spine, and is inserted along the dorsal midline between the base of the anal fin spine and the pelvic fin spines. The ornamentation on the dorsal fin spine is identical to that of the other fin spines, with a smooth leading edge rib and a posterior field of smooth, fine longitudinal striations ( Fig. 6 View FIG A-C). The dorsal fin spine is supported by a thin, ossified, rhombic basal plate ( Fig. 6C View FIG ).
The base of the dorsal fin spine is covered by large smooth-crowned scales, and there is an abrupt transition in scale size from the body to the basal portions of the dorsal fin web ( Fig. 6 View FIG A-C). The smooth-crowned dorsal fin scales are aligned in rows, and scales decrease in size towards the fin margin. Fin scales have narrow crowns with acutely pointed posterior tips, correspondingly little basal tissue, and a low neck in comparison to typical body scales. The dorsal fin web is triangular and likely reached the tip of the dorsal fin spine ( Figs 2 View FIG ; 3 View FIG ), although the exact margin of the fin cannot be confirmed. None of the specimens of P. eppleri n. gen., n. sp. have a complete dorsal fin web.
Prepelvic spines and pelvic girdle
A single pair of prepelvic spines is visible on UALVP 41860, 42652 and 42653, and these spines are positioned closer to the pelvic girdle than to the pectoral girdle ( Figs 2 View FIG ; 3 View FIG ). The prepelvic spines are short, have a broad open basal cavity with a shallow insertion in the body wall, and are ornamented with fine striations.
The pelvic fin spines are slender, shallowly inserted, have similar ornamentation as the dorsal, pectoral, and anal fin spines ( Figs 2 View FIG ; 3 View FIG ), and are positioned anterior to the base of the dorsal fin spine. Pelvic fin spines support a large fin web, but the outline of the fin cannot be determined in the available specimens. The base of the pelvic fins extends posteriorly to near the origin of the anal fin spine. Smooth-crowned scales on the pelvic fin web are similar in size and shape to those on the dorsal fin.
Anal fin and spine
An anal fin and fin spine are positioned just posterior to the dorsal fin spine origin ( Figs 2 View FIG ; 3 View FIG ). The anal fin spine is long, slender, shallowly inserted, and curves posteriorly near the tip, and if depressed, would not contact the origin of the hypochordal lobe of the caudal fin. The ornamentation of the anal fin spine is identical to that of the dorsal fin spine. The anal fin web is broad-based, terminates just anterior to the origin of the caudal fin, and its smooth-crowned scales are similar in size and shape to those on the dorsal fin.
Caudal fin
The caudal peduncle is deep and tapers posteriorly along the caudal fin axis ( Figs 2 View FIG ; 3 View FIG ). The axis of the caudal fin is deflected dorsally and supports a large hypochordal fin web. The scales of the axis of the caudal fin have narrow, elongate, tear-drop-shaped crowns, with rhombic shaped bases ( Fig. 6D, E View FIG ). These scales decrease in size towards the posterior tip of the caudal fin axis.
The caudal fin web has a straight to concave trailing margin, and the caudal fin scales are aligned in rows ( Figs 3 View FIG ; 6F View FIG ). Each fin scale is narrow, has a round anterior margin, and an acutely pointed trailing tip, and fin scales decrease in size towards the fin margin. The caudal fin web does not reach the tip of the caudal fin axis, and a low epichordal lobe is present ( Figs 2 View FIG ; 3 View FIG ); the scales on the epichordal lobe are smaller than those on the caudal peduncle.
Scales
There is an abrupt transition between head scales and typical body scales dorsal to the branchial chamber ( Figs 3 View FIG ; 5E View FIG ). Body scales behind the head are small and are comparable in size to scales on the basal portions of the fin webs. The crown of each body scale is smooth and flat, with a rounded anterior margin and an acutely pointed posterior apex ( Fig. 7 View FIG ). All body scales are aligned in oblique rows, and the posterior apex of each scale overlaps the anterior margin of scales in the next posterior row. The largest body scales are found around the base of the dorsal fin and on the caudal peduncle ( Fig. 6A, D View FIG ).
The neck of each body scale is developed as a cone that surrounds the mass of basal tissue ( Fig. 7 View FIG E-I). The scale neck is attached to the anterior two-thirds of the scale crown, but the narrow diameter neck canal pores which link to form radial canals inside each scale have not been located ( Fig. 7 View FIG E-G). Body scales have tumid bases ( Fig. 7C View FIG , E-I), and in ventral view, body scales have either a round, rhombic, or oval mass of basal tissue deposited within the rim of neck tissue. There is a well-developed horizontal flange that marks the junction between the neck and the base ( Fig. 7 View FIG E-I). The thickest part of the scale base is centered, or positioned just anterior of center, relative to the periphery of the basal tissue.
Thin sections of body scales show a typical acanthodid-type microstructure ( Fig. 7H, I View FIG ). Thin sections show that the scale primordium is small relative to the rest of the crown, and is covered with four or five thin growth zones of superpositioned odontodes. Details of dentine tubules and ascending canals could not be determined given poor scale preservation at the MOTH locality. Radial canals exit scales through narrow neck canal pores, just above the neck-base junction. Basal tissue appears to be acellular and shows traces of Sharpey’s fibres.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Promesacanthus eppleri
| Hanke, Gavin F. 2008 |
Acanthodiformes
| HANKE G. F. 2002: 1072 |
Mesacanthidae
| WILSON M. V. H. & HANKE G. F. & SOEHN K. L. 2000: 139 |