
Polydorella kamakamai, Williams, 2004
|
publication ID |
https://doi.org/10.1080/0022293031000155395 |
|
persistent identifier |
https://treatment.plazi.org/id/537A7F7A-FF94-FD2B-564E-9B71707DFBE9 |
|
treatment provided by |
Felipe |
|
scientific name |
Polydorella kamakamai |
| status |
sp. nov. |
Polydorella kamakamai View in CoL , sp. n.
( figures 1–5 View FIG View FIG View FIG View FIG View FIG )
HOLOTYPE: Philippines, Puerto Galera , Agus Beach, y 3 m, 1 July 2000, from Clathria (Thalysias) cervicornis (Thiele, 1903) ( USNM 1008710 About USNM ).
PARATYPES: same data as for holotype ( USNM 1008711 About USNM : 270z paratypes; USNM 1008712 About USNM : 10 paratypes, on nine SEM stubs). Philippines, Puerto Galera, Small La Laguna Beach , y 3 m, 24 July 2000, from Clathria (Thalysias) cervicornis ( AMNH 4347 About AMNH : 180z paratypes) .
Etymology. The specific name kamakamai is derived from the Filipino word for supernatural beings that are small, human-like creatures possessing long beards and nails used to pinch offending children ( Jocano, 1969), reminiscent of the acicular neurosetae of anterior segments in this species.
Description
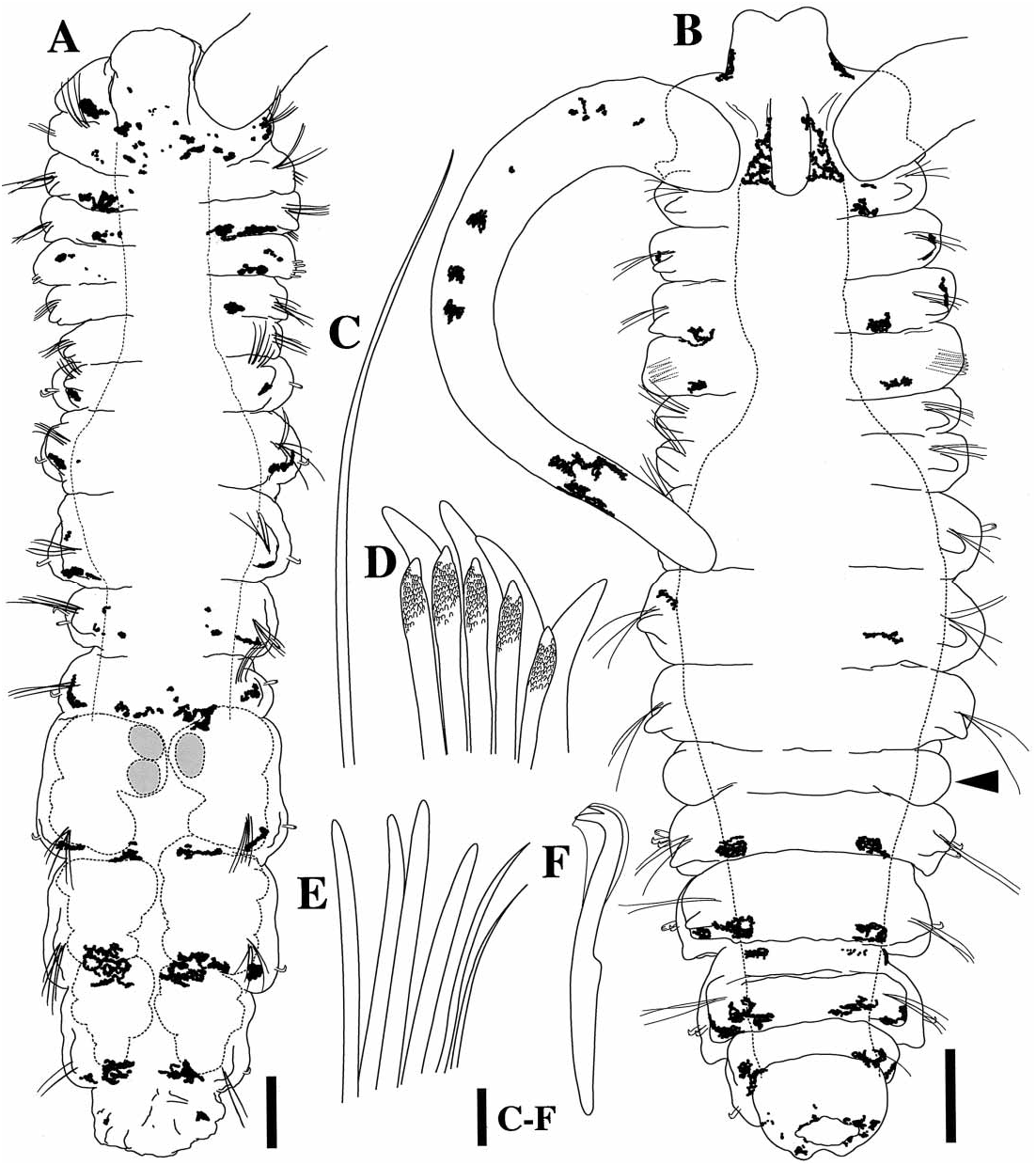
Holotype. 1.3 mm long, 0.3 mm wide at segment 7; 14 segments. Prostomium bifid; caruncle short, terminating between posterior margin of segment 1 and middle of segment 2; occipital tentacle and eyes absent ( figures 1–3 View FIG View FIG View FIG ). Palps extend posteriorly for 10–12 segments. Black pigmentation in irregular patches on palps, sides of peristomium, prostomium, along caruncle, dorsal and ventral sides of all segments and pygidium ( figures 1 View FIG , 3 View FIG ).
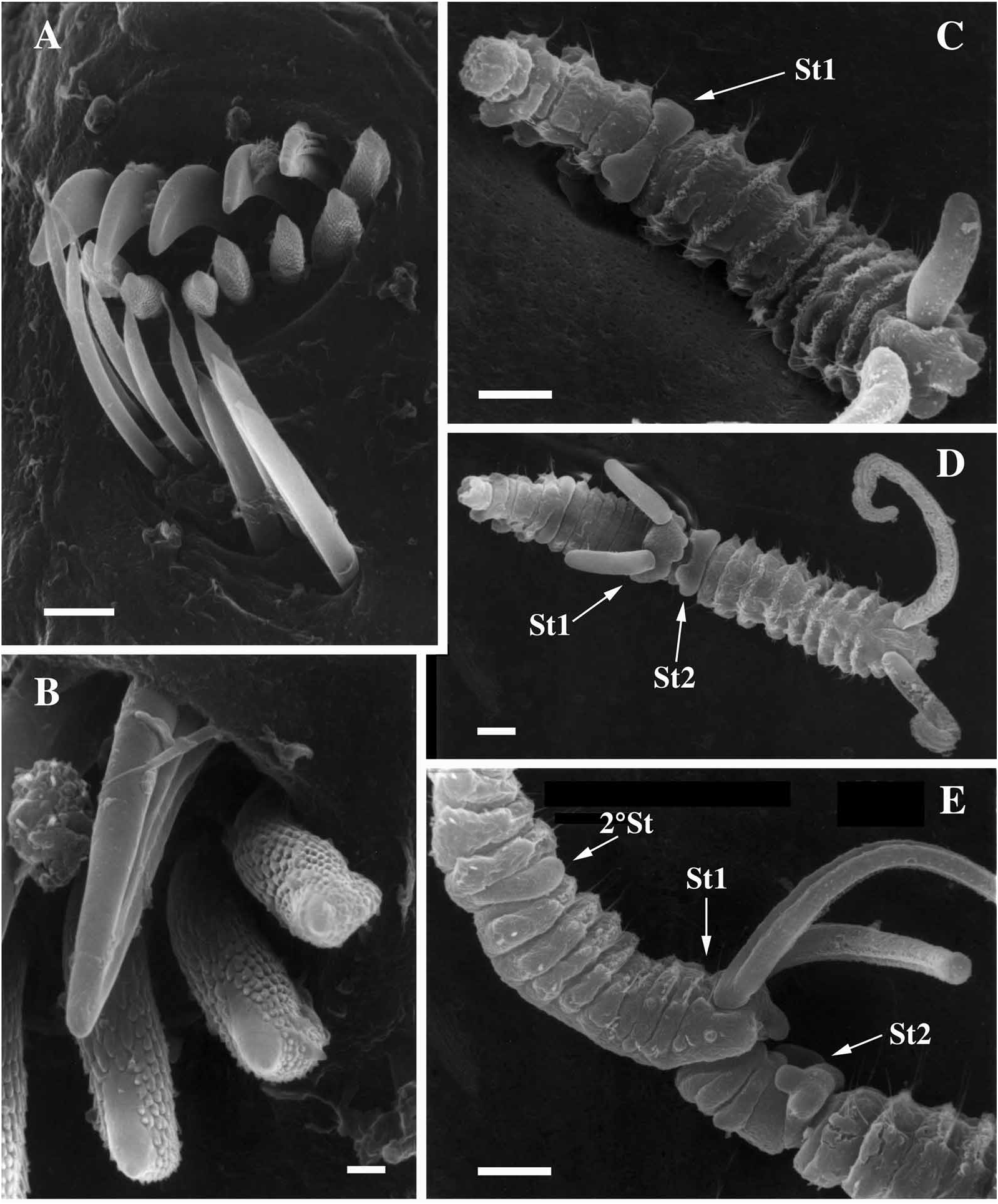
Segment 1 without notopodial lobes or notosetae; neuropodia lobes present and neurosetae absent. Capillary notosetae of segments 2–4, 6 and subsequent segments in two rows; no specialized posterior notosetae. Neurosetae of segments 2–5 of two types, arranged in two rows; dorsal row composed of up to three acicular spines on segment four, ventral row composed of up to four capillaries on segment four ( figure 1C, E View FIG ); segments 6 and 7 with reduced number of acicular spines and capillaries; three bidentate hooded hooks begin on segment 8, not accompanied by capillaries; hooks with approximately right angle between main fang and shaft, narrow angle between main fang and apical tooth, with constriction on shaft ( figure 1F View FIG ).
Segment 5 slightly modified, about the same size as segments 4 and 6, with acicular spines and capillary neurosetae in same arrangement as on segments 2–4; notosetae absent ( figure 2A View FIG ). Two types of major spines on segment 5 in approximately straight, horizontal rows; ventral row of four acicular spines with small digitiform bosses ( y 0.3 m m) closely applied to the apical ends of the spines, lacking on tips of the spines ( figures 1D View FIG , 2A, B View FIG ), and a dorsal row of four falcate spines ( figures 1D View FIG , 2A, B View FIG ).
Branchiae absent. Dorsal ciliary bands apparent from segment 3 continuing to terminal segments of stock individual ( figure 2C–E View FIG ).
Narrow pharynx to segment 7 leading to broad intestine, narrowing in posterior segments; without gizzard-like structure in digestive tract.
Pygidium reduced, knob-shaped ( figure 1B View FIG ); with large cells ( y 30 m m) within pygidium and terminal segments ( figure 3 View FIG ).
Variability. All specimens examined exhibited black pigmentation on the palps, peristomium, prostomium, surrounding the caruncle, sides of anterior and posterior segments, and on the pygidium. Pigmentation was most prominent on the dorsal side but was also present on the ventral side of segments. A bifid prostomium and caruncle extending to middle of segment two was observed in
all specimens; no specimens possessed branchiae. Acicular neurosetae were found in approximately vertical rows reaching four in number on segment 5, accompanied by a dorsal row of typically four to six capillary neurosetae. The number of acicular neurosetae was reduced to two or three in segments 6 and 7, accompanied by one or two capillary neurosetae.
The fifth segment contained four to seven major spines in both the dorsal and ventral rows; 20% of the specimens examined ( N ~50) possessed one more additional spine in the ventral row than the dorsal row; all other specimens possessed equivalent numbers of spines in both rows. Hooded hooks numbered three or four on segment 8, reaching a maximum of three to five hooded hooks on
segments 8–11. The pygidium was variable, from a reduced, knob-shape ( figures 1B View FIG , 2C, D View FIG ) to a more elongate conical shape ( figure 3 View FIG ); small bumps were often observed surrounding the anus.
Distribution Philippines: Oriental Mindoro. Remarks
Polydorella kamakamai is unique within the genus in possession of acicular neurosetae in segments 2–7 and the fifth segment containing ventral row of major spines with digitiform bosses. The species is further differentiated from P. dawydoffi , P. prolifera and P. stolonifera by the lack of branchiae.
Ecology
Polydorella kamakamai was found to construct small mud tubes (up to 5 mm) on the surface of the sponge, Clathria (Thalysias) cervicornis . The worms were found in fairly high abundance, with approximately 280 worms on a sponge consisting of eight main branches approximately 100 mm long and 7 mm in diameter ( y 1892.0 cm 2). Some specimens contained material in the gut the same colour as the preserved sponge; additional behavioural and histological studies are required to determine if the worms feed on the tissue of sponges as has been documented in some other polychaetes (see Martín and Britayev, 1998). Many tubes contained two individuals with dorsal sides apposed and anterior ends facing in opposite directions; tubes containing greater than two individuals were not observed. When disturbed, worms were able to move quickly out of their tubes and efficiently crawl across the surface of the sponge.
Asexual reproduction
Most specimens examined ( N ~290) exhibited some stage of asexual reproduction via paratomy ( figure 4 View FIG ). Single parental or stock individuals consist of 13–15 segments ( figure 5A View FIG ). Stock individuals then exhibit a growth zone which follows segment 10 ( figures 1B View FIG , 2C View FIG , 5B View FIG ). The growth zone is distinct from anterior and posterior segments in its opaque colouring and smooth form. The opaque colouring of the growth zone may come from the breakdown of muscular material as the zone prepares for the formation of a new stolon body ( Schroeder and Hermans, 1975). The growth zone exhibits two sections: an anterior section that will become the new segment 11 (denoted as 11 ’, following Radashevsky (1996) where ’ indicates newly developed stock segments) and the posterior portion which produces the developing stolon 1.
After further development of the growth zone, palp buds are formed and the new stock segment anterior to the head of the developing stolon 1 exhibits setae. New segments of the stolon body then appear to form simultaneously (overt distinction of 10 segments displayed at once from subdivision of the elongated growth zone). Ultimately, stolon 1 consists of 10 new anterior segments and three to four posterior segments derived from the stock individual ( figure 5D View FIG ). Stolon 2 is formed following 11 ’. Prior to complete development of the first 10 segments of stolon 1 (i.e. lacking externally observable setae), a secondary growth zone is formed following segment 10. Formation of stolon 2 proceeds as described above and the secondary growth zone of stolon 1 then exhibits palp buds ( figures 2D, E View FIG , 5D View FIG ). A third stolon (stolon 3) is observed in some individuals behind 11 ’. In this way, a chain of five developing individuals is formed (stock, stolon 3, stolon 2, stolon 1 and the secondary stolon) ( figures 3A, B View FIG , 5E View FIG ). The process of asexual reproduction is shown in figure 5 View FIG .
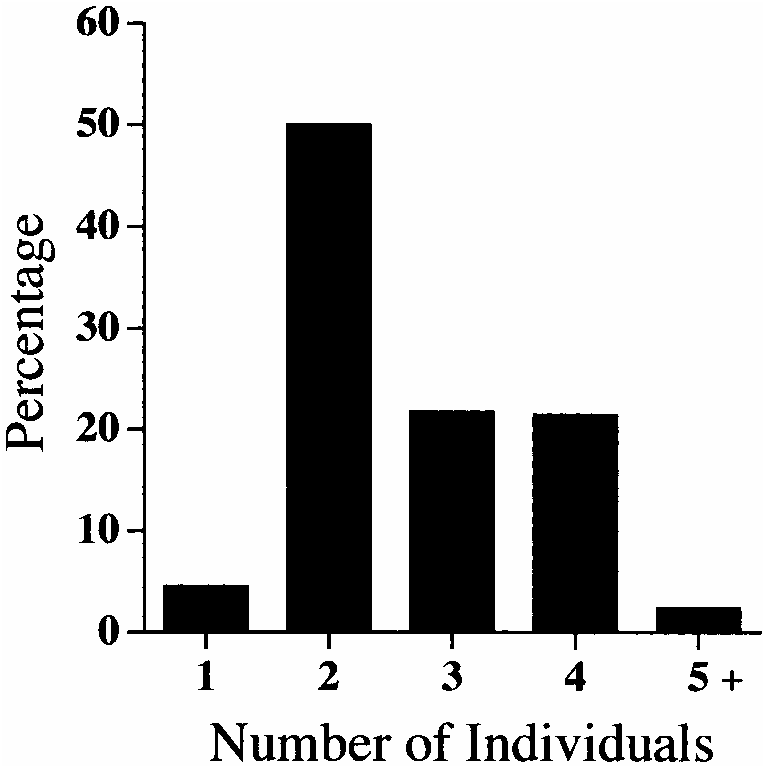
It appears that in most cases paratomic division takes place when four or five individuals are present in a chain. Stolon 1 (with secondary stolon) breaks from the stock individual (stolon 3 and stolon 2) ( figure 5E View FIG ). Prior to division, the posterior segments of stolon 2 are found in a wedge-shape under the head of stolon 1 ( figures 2E View FIG , 3B View FIG ). At this time, palps of stolon 1 are approximately one-half to threequarters the length of stock individuals. Following division, the pygidium is fully formed on the posterior end of stolon 2. Stolon 1 (with secondary stolon) and stock (with stolon 3 and stolon 2) individuals then repeat the course of asexual reproduction described above. Following division, the worms apparently do not immediately leave their original tubes and construct new ones, as evidenced by tubes containing two individuals. The division of paratomic chains with stock individuals lacking a stolon 3 or stolon 2 leads to the presence of single individuals within the population. In only seven (2.4%) cases were the stock individuals within paratomic chains observed to possess 13–15 stock segments prior to division; this low number explains the relative rarity of single individuals lacking growth zones ( figure 4 View FIG ).
The colonies were dominated by specimens composed of chains of two individuals (sometimes with developing growth zones); chains of three and four individuals were approximately half as abundant as chains of two individuals ( figure 4 View FIG ). Only rarely were single individuals and chains of five individuals or greater observed ( figure 4 View FIG ).
Sexual reproduction
In addition to asexual reproduction, one specimen composed of 15 segments was found with eggs ( figure 1A View FIG ). Segments 10–12 of the specimen were narrower than preceding segments (no growth zone was exhibited), while segments 13–15 were expanded and contained yolky eggs. Eggs were oval in shape; 43.3 m m (¡5.8) by 62.2 m m (¡6.8) ( N ~11). Based on the volume of the eggs within this specimen, segments 13–15 are estimated to contain 36–42, 26–34 and 15– 20 eggs, respectively.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.