
Pinnixa banzu, Komai, Tomoyuki, Nishi, Eijiroh & Taru, Masanori, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3793.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:99154563-6A9C-49C3-AEF1-1A54D9A60C7A |
|
DOI |
https://doi.org/10.5281/zenodo.6131951 |
|
persistent identifier |
https://treatment.plazi.org/id/03DA87E4-7120-FFFE-FF12-4CA0FEC49E1D |
|
treatment provided by |
Plazi |
|
scientific name |
Pinnixa banzu |
| status |
sp. nov. |
Pinnixa banzu View in CoL n. sp.
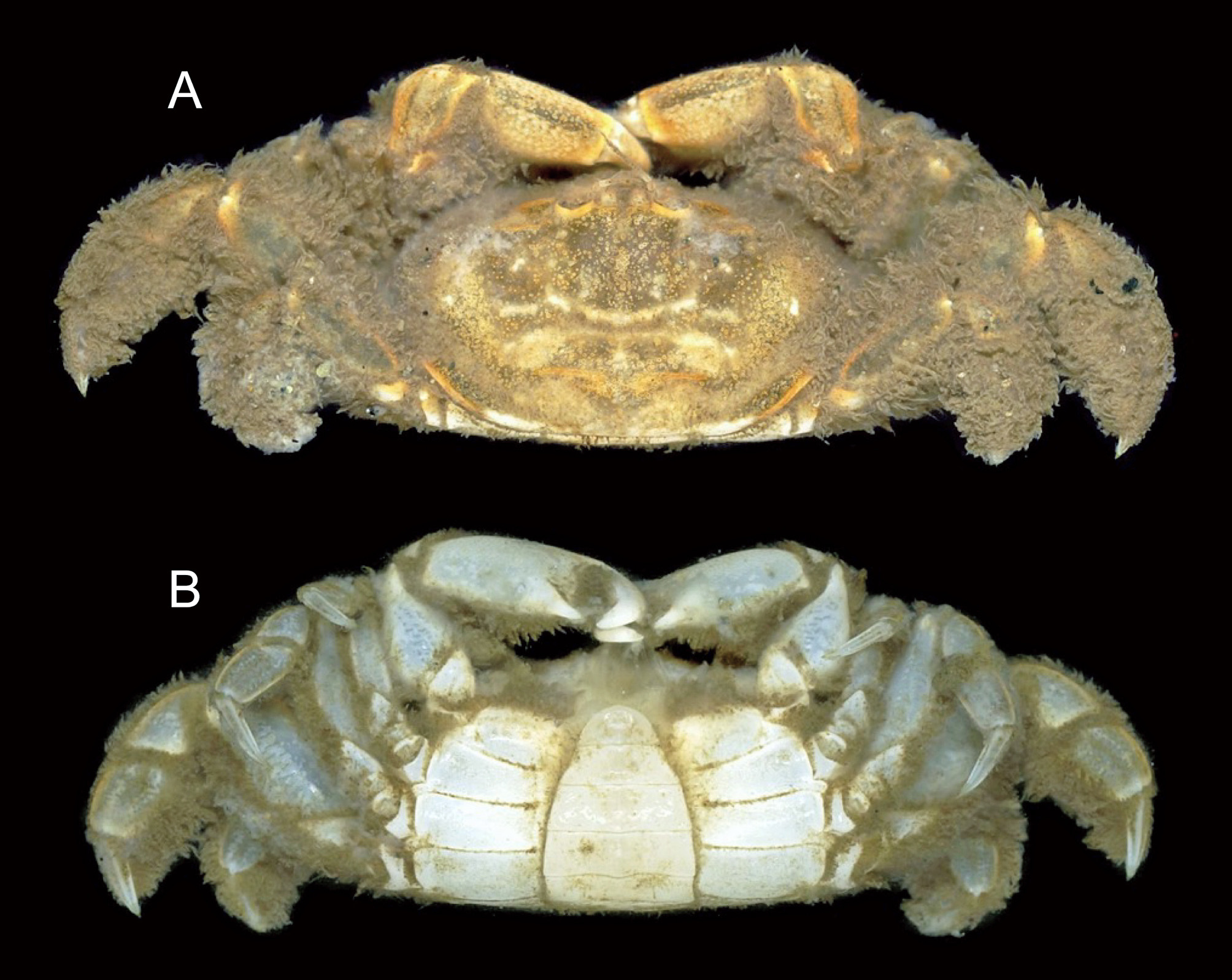
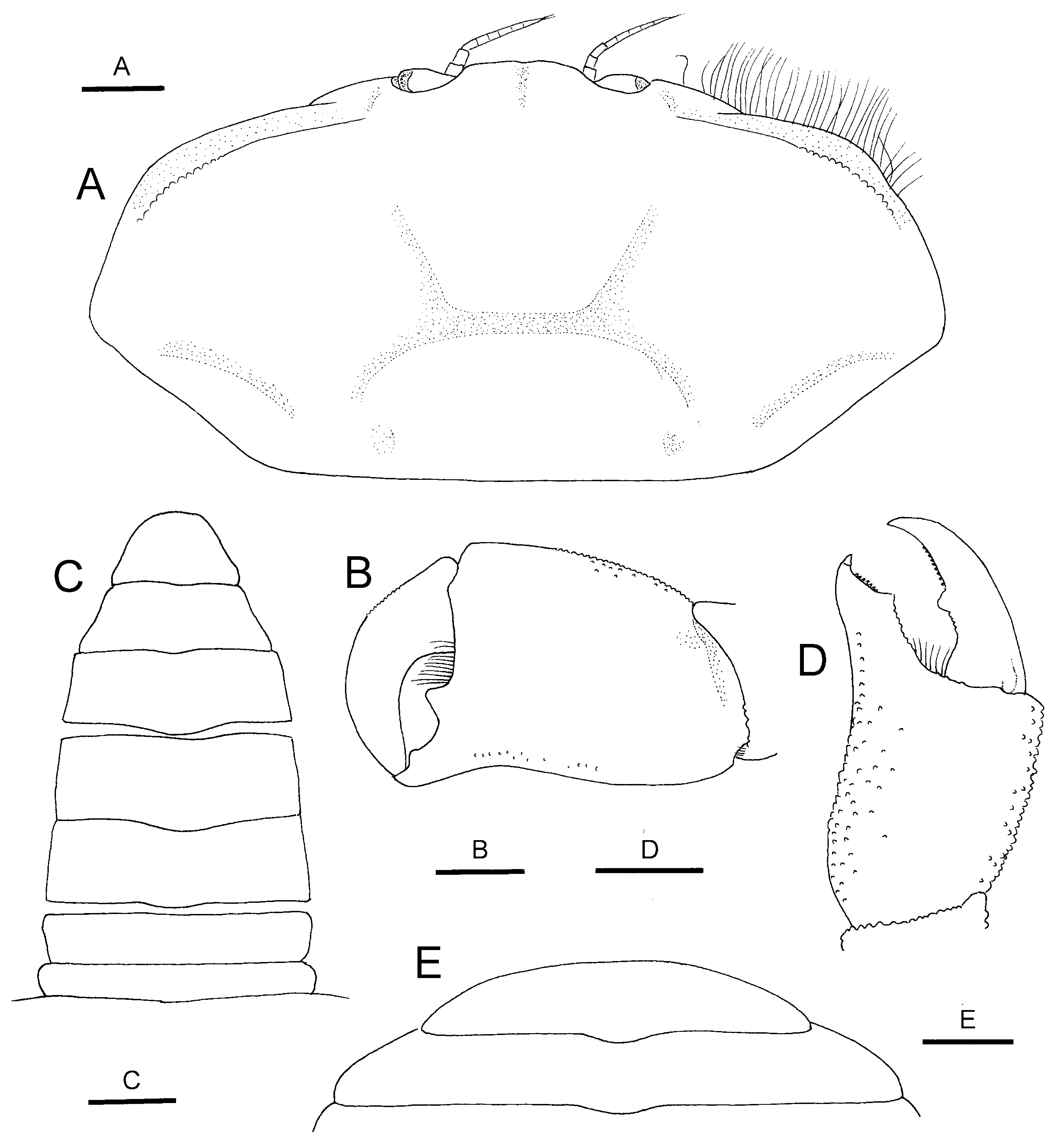
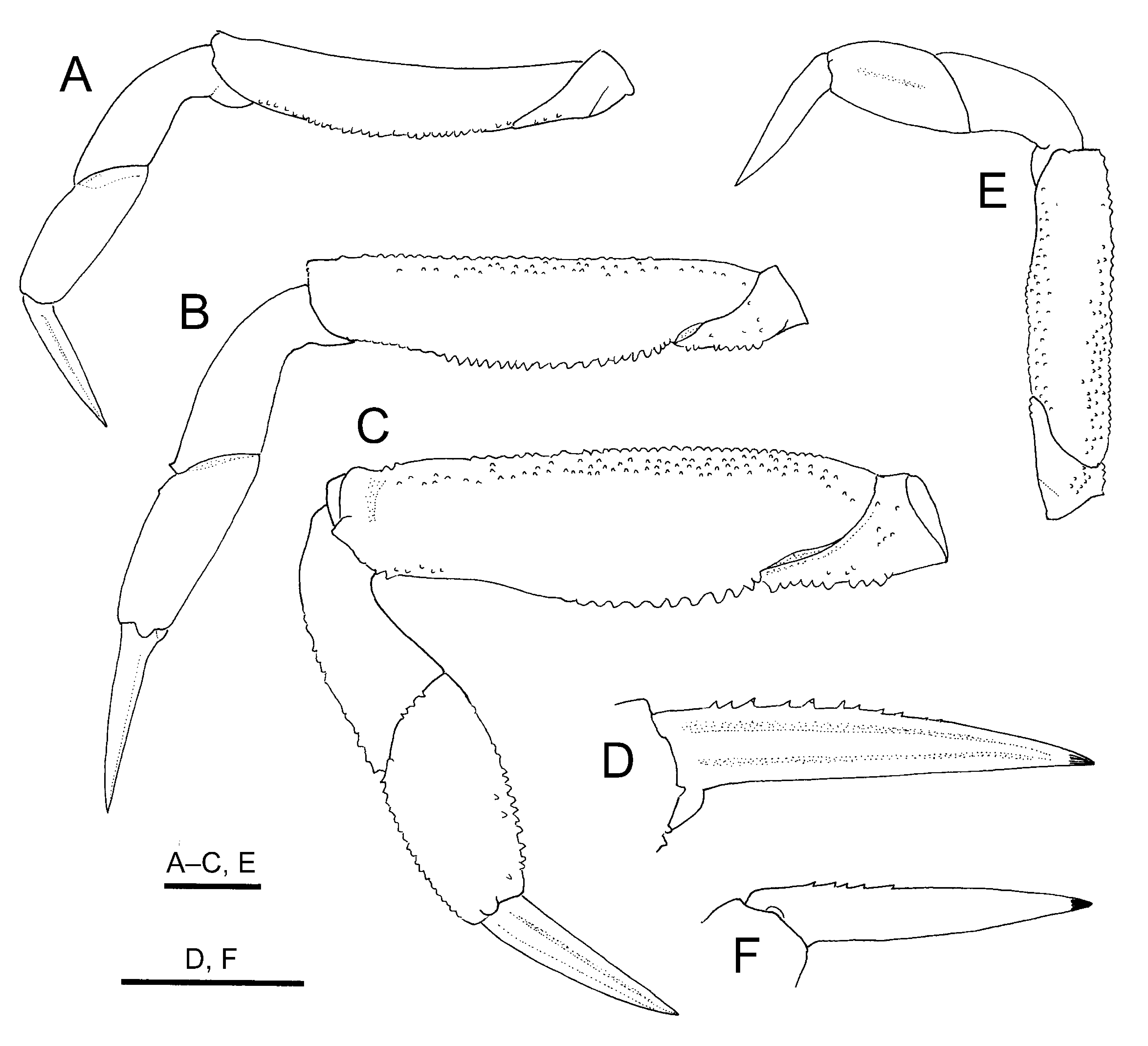
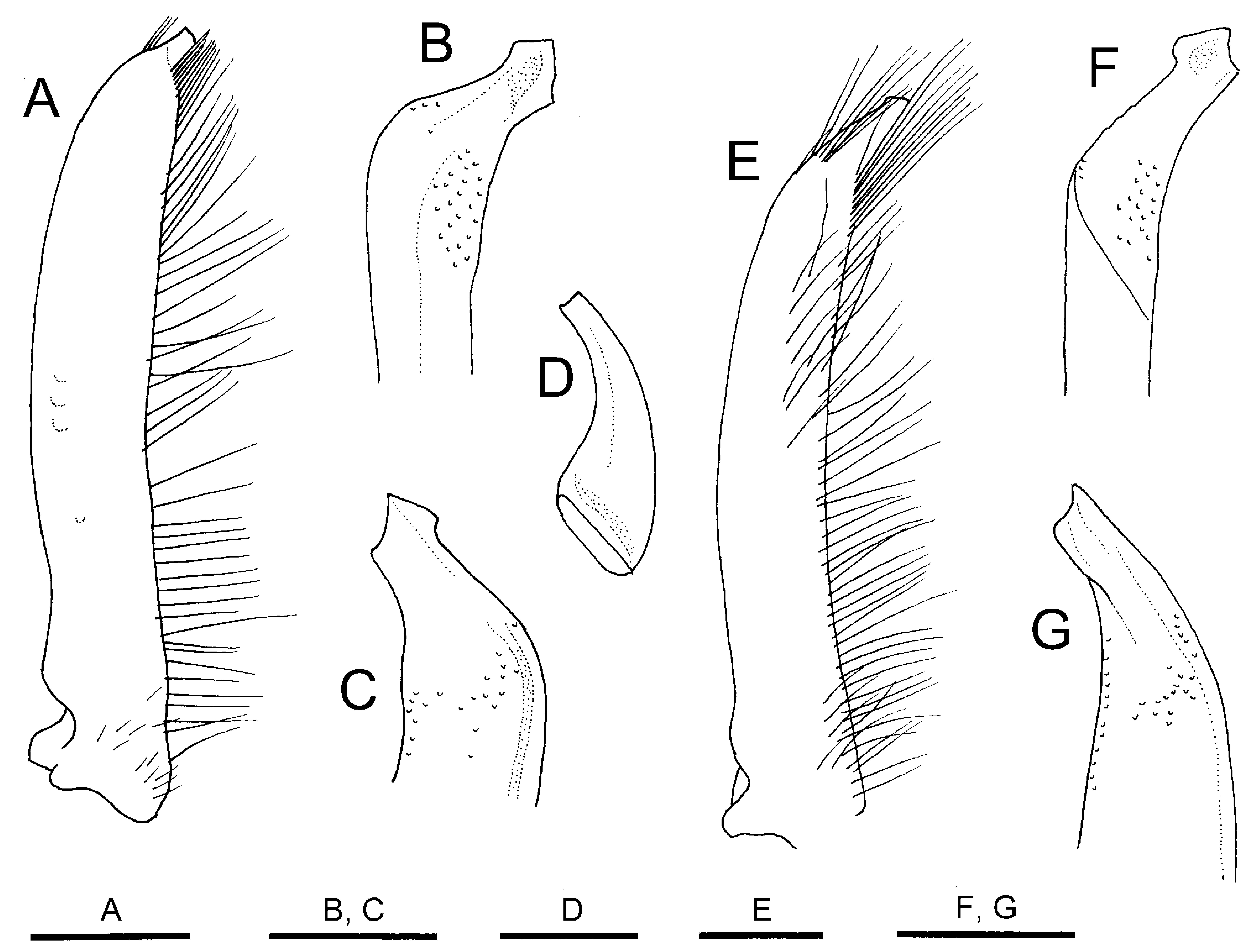
( Figs 2 View FIGURE 2 , 3 View FIGURE 3 , 4 View FIGURE 4 , 5A‒D View FIGURE 5. A – D , 6 View FIGURE 6 , 7 View FIGURE 7 )
Material examined. Holotype: Banzu , estuary of Obitsu River, Kisarazu, Chiba Prefecture, Tokyo Bay, 25 June 2010, associated with Chaetopterus cautus , coll. E. Nishi, male (6.6 × 13.1 mm), CBM-ZC 12293.
Paratype: same data as holotype, 1 female (5.8 × 11.6 mm), CBM-ZC 12294; same locality as holotype, 1 June 2012, associated with C. cautus , coll. M. Taru, 1 female (5.7 × 11.2 mm), CBM-ZC 12295.
Other material: same locality as holotype, 21 April 2011, coll. E. Nishi, 1 pre-adult female (4.1 × 7.1 mm), CBM-ZC 12296.
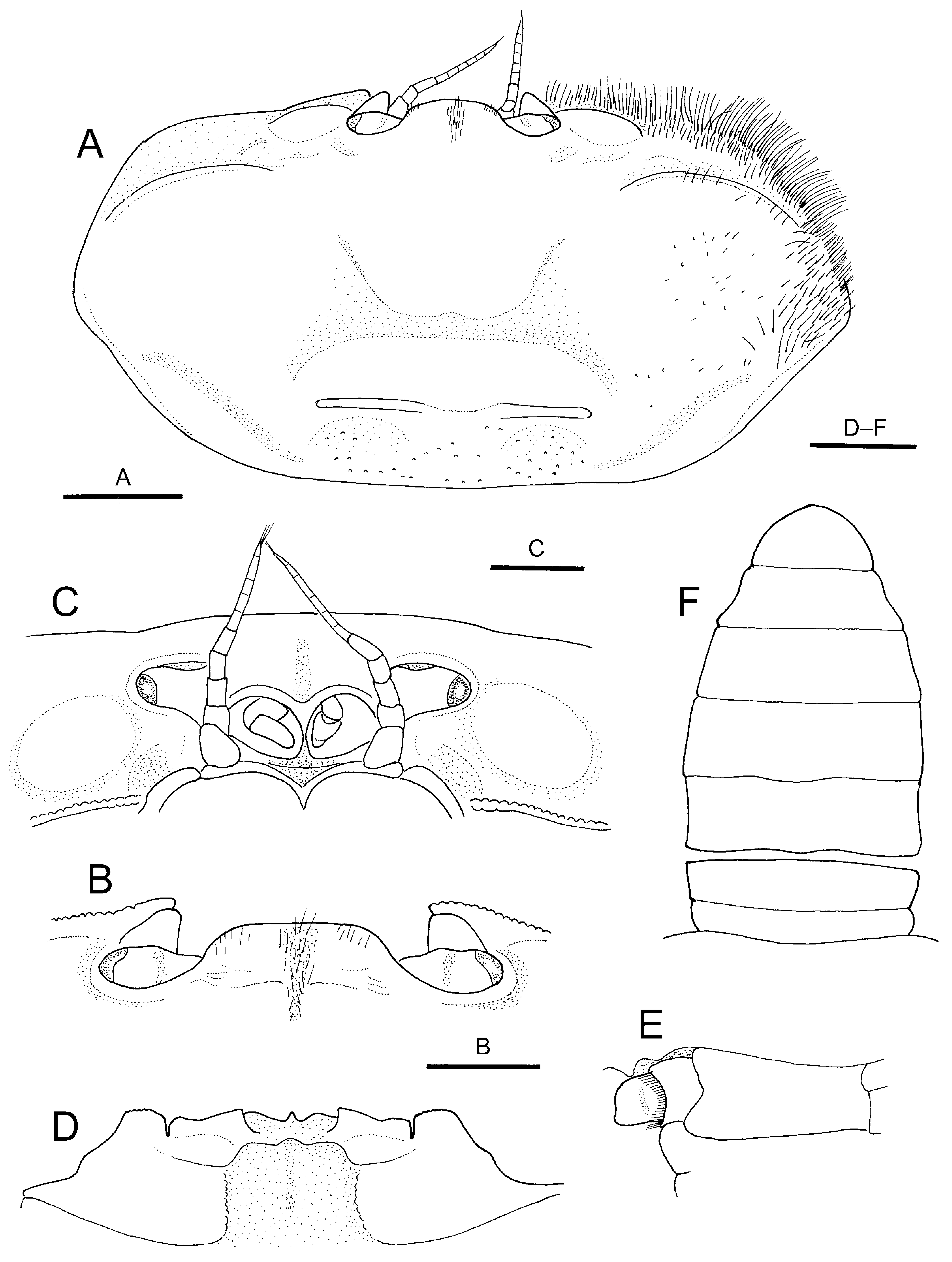
Description. Adult male (holotype). Carapace ( Figs 2 View FIGURE 2 A; 3A) about 2.0 times wider than long. Front ( Fig. 3 View FIGURE 3 B, C) slightly deflexed, with faint median sulcus, width slightly more than 0.1 of carapace width; frontal margin merging into inter-antennular septum. Dorsal surface of carapace microscopically rugose, slightly convex transversely, no distinct grooves demarcating regions, with scattered minute pits, numerous short setae laterally; gastric region glabrous, slightly convex; branchial regions slightly convex, sloping to lateral side; gastro-cardiac sulcus moderately deep; cardiac region somewhat elevated; cardiac ridge straight, low, but clearly delimited, not extending laterally beyond cardiac region, widely interrupted medially; pair of shallow depressions present posterior to cardiac region; posterior part of cardiac, intestinal regions fairly sloping posteriorly; branchial ridge (delimiting anterolateral margin) low, minutely granular (granules becoming conspicuous laterally), extending posterolaterally from orbit to lateral branchial region, not reaching junction between anterolateral ventral, posterolateral margins. Subbranchial regions fairly advanced; anterolateral ventral margin slightly sinuous in dorsal view, bordered by low, minutely granular ridge extending from anterolateral angle of buccal cavity, fringed with short plumose setae; junction between anterolateral ventral, posterolateral margins rounded; posterolateral margin smooth, slightly elevated; posterior margin straight.
Epistome ( Fig. 3 View FIGURE 3 C) subtriangular, very short, somewhat depressed below. Anterior margin of buccal frame rounded in either side of sharp endostomal ridge.
Orbit ( Fig. 3 View FIGURE 3 B, C) smooth, entire, about 0.6 frontal width, completely filled by eye; inner orbital angle slightly produced. Pterygostomial region ventrolateral to orbit slightly inflated.
Ocular peduncles ( Fig. 3 View FIGURE 3 B, C) short, stout, slightly constricted medially, transversely retracted in orbit; corneas small, darkly pigmented.
Antennule ( Fig. 3 View FIGURE 3 C) obliquely transversely folded in antennular fossa; antennular fossa slightly smaller than orbit.
Antenna ( Fig. 3 View FIGURE 3 C) closing orbit with second segment of peduncle; basal segment suboval; second segment slightly movable; third segment freely articulated.
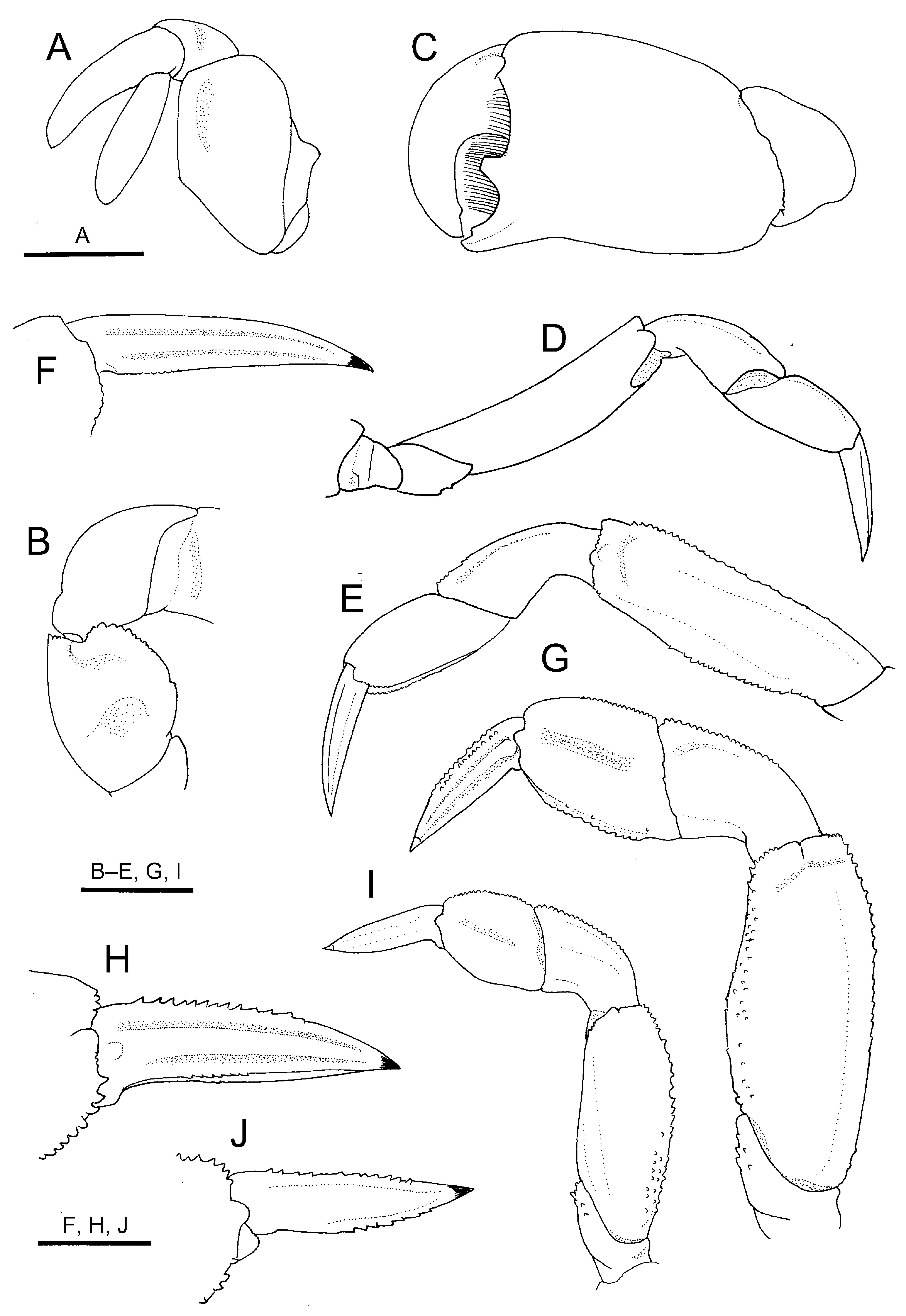
Third maxilliped ( Fig. 4 View FIGURE 4 A) with conspicuously short basis. Ischium completely fused with merus. Ischiummerus fused segment subtrapezoidal; outer surface smooth, with faint longitudinal sulcus adjacent to mesial margin. Carpus cup-shaped, articulated at distal margin of merus. Propodus, dactylus bearing long setae distally, both elongated, spatulate in shape; dactylus articulated at proximal 0.2 of propodus, distinctly overreaching tip of propodus. Exopod with prominent lobe at midlength of lateral margin; flagellum slender, consisting of 2 articles, distal article longer than proximal article, bearing long terminal setae.
Chelipeds ( Figs 2 View FIGURE 2 A, B; 4B, C) symmetrical. Merus trigonal in cross section; dorsal surface with low protuberance at midlength, distal half obscured by numerous setae, dorsodistal margin minutely granulate, fairly upturned; inner dorsal surface convex, granular: outer (posterior) surface slightly convex, with patches of setae; ventral surface convex transversely, glabrous. Carpus unarmed, with setae on inner distal margins. Chela compressed, with plumose setae on proximal to ventral parts of inner surface, cutting edge of fixed finger, hiatus between fingers filled by dense plumose setae. Palm subrectangular, slightly longer than wide, glabrous on dorsal, outer, inner surfaces; dorsal, ventral surfaces rounded; distal margin of palm to cutting edge of fixed finger nearly perpendicular. Fixed finger deflexed, slightly curved distally; cutting edge nearly transverse, armed with prominent, distally truncate tooth proximal to midlength, minute subterminal tooth. Dactylus strongly curved, strongly deflexed, sickle-shaped, terminating in subacute tip; dorsal surface rounded; cutting edge with minute subdistal denticle.
Ambulatory legs (second to fifth pereopods) ( Figs 2 View FIGURE 2 A, B; 4D, E, G, I) dissimilar in shape, length, relative length (third=fourth>second>fifth); extensor, flexor, dorsal surfaces of meri to propodi with thick plumose setae, ventral surfaces nearly glabrous.
Second pereopod ( Fig. 4 View FIGURE 4 D) moderately slender, 1.4 of carapace length. Merus slightly recurved; extensor surface rounded, not carinate; flexor margin with row of granules. Carpus slightly carinate on extensor margin. Propodus somewhat compressed, oval in cross section, 1.4 times longer than wide; extensor margin slightly carinate; flexor margin faintly carinate; flexor distal margin minutely granular. Dactylus slightly curved, subconical, about 1.4 propodus length, with 5 longitudinal carinae (1 extensor, 1 dorsal, 1 ventral, 2 flexor), flexor carina with row of granules in proximal 0.4.
Third pereopod ( Fig. 4 View FIGURE 4 E) 1.7 carapace length. Merus suboval in cross section; extensor surface rounded, not carinate, with row of minute granules; flexor margin with row of granules. Carpus bluntly carinate on extensor margin, with row of sharp granules in distal 0.3. Propodus 1.3 times longer than wide; extensor margin bluntly carinate, unarmed; dorsal surface with shallow median sulcus; flexor surface with 2 rows of granules (dorsal row in distal 0.2, ventral row in distal half). Dactylus ( Fig. 4 View FIGURE 4 F) nearly straight, 1.4 propodus length, with 5 longitudinal carinae; extensor and flexor carinae with sharp granules proximally.
Fourth pereopod ( Fig. 4 View FIGURE 4 G) fairly stout, compressed, 1.7 carapace length. Merus suboval in cross section, 2.4 times longer than wide; extensor surface generally rounded, not carinate, with 2 rows of sharp granules increasing in size distally; dorsal and ventral surfaces smooth; flexor surface with sparse granules. Carpus bluntly carinate on extensor margin, with row of minute granules over entire length; dorsal side of distal margin also with granules. Propodus approximately as long as wide; extensor margin bluntly carinate, with row of minute granules in proximal half; dorsal surface with shallow median sulcus; flexor surface flat, dorsal side with few granules distally, ventral side sharply carinate, serrated with row of sharp granules. Dactylus ( Fig. 4 View FIGURE 4 H) nearly straight, 1.3 propodus length, about 3.7 times as long as wide, with 6 longitudinal carinae (2 extensor, 1 dorsal, 1 ventral, 2 flexor), extensor and flexor carinae each with row of granules.
Fifth pereopod ( Fig. 4 View FIGURE 4 I) 1.3 carapace length, distinctly overreaching distal margin of merus when extended laterally. Merus suboval in cross section, about 2.7 times longer than wide; extensor margin with row of minute granules; dorsal and ventral surfaces smooth; flexor surface with ventral margin distinctly delimited, bearing row of granules. Carpus bluntly carinate on extensor margin, with row of minute granules on distal 0.7. Propodus approximately as long as wide, with row of granules on extensor margin; dorsal surface with shallow median sulcus; flexor margin convex in outline, without carinae. Dactylus ( Fig. 4 View FIGURE 4 J) nearly straight in dorsal view, slightly upturned in extensor view, 1.4 length of propodus, armed with rows of sharp, corneous-tipped granules or denticles on ventral side of extensor and flexor margins.
Thoracic sternum ( Fig. 2 View FIGURE 2 B) broad. Sternites 1–3 ( Fig. 3 View FIGURE 3 D) depressed, continuing to sterno-abdominal cavity. Sternite 1 with convexity on either side of small median tooth on anterior margin. Suture between sternite 2/3 clearly demarcated; other sutures between sternites interrupted medially. Short, narrow slit dividing sternite 3/4 just lateral to base of third maxilliped. Sternites 4–8 smooth, glabrous; exposed part of sternite 8 ( Fig. 3 View FIGURE 3 E) subrectangular, perpendicular to horizontal plane of carapace, lateral margin shallowly notched.
Abdomen ( Fig. 3 View FIGURE 3 F) of free 6 somites, widest at somite 4, about 1.8 times longer than wide, occupying about one-third width of thoracic sternum, slightly narrowed distally to somite 3, lateral margins fringed with short sparse setae. Somites 1–3 equal in width, somite 1 about half length of somite 2; somites 3–6 subequal in length, oblong (somite 3) or subtrapezoidal (somites 4–6). Somite 3 with slightly concave lateral margins. Somite 4 about 3.3 times wider than long. Somite 6 subtrapezoidal, with concave lateral margins. Telson subsemicircular, about twice wider than long, subequal in length to somite 6.
First gonopod ( Fig. 5A View FIGURE 5. A – D ) strongly flattened, moderately slender, gently curved laterally, reaching suture between sternites 3/4; terminal process ( Fig. 5B, C View FIGURE 5. A – D ) relatively stout, directed distolaterally, chitinous distally, flared distally, with shoulder on outer margin; apex obliquely truncate. Second gonopod ( Fig. 5D View FIGURE 5. A – D ) short, curved dorsally, tapering distally to truncate apex.
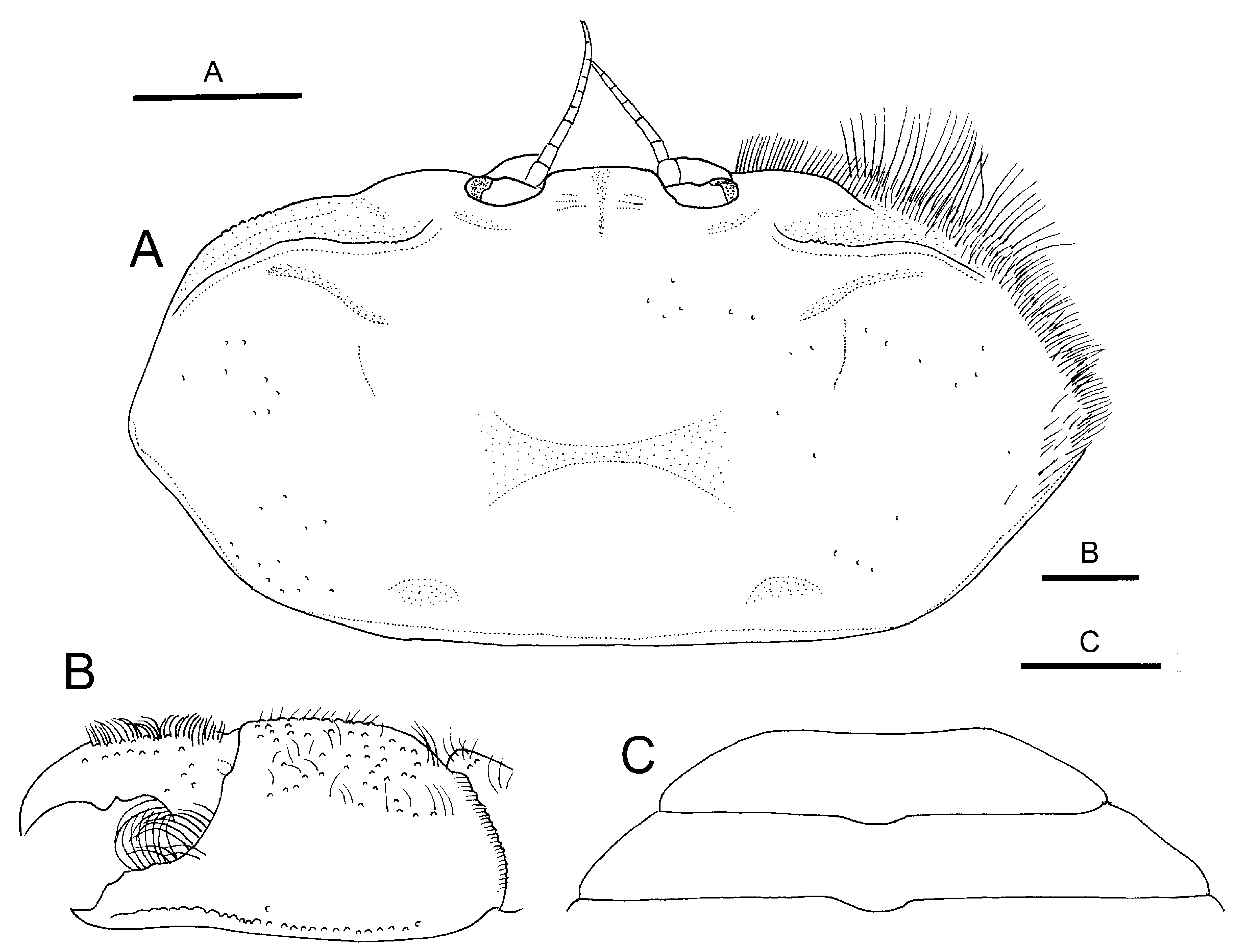
Adult females (paratypes). Carapace ( Figs 6 View FIGURE 6 A; 7A) about 2.0 times as wide as long (ratio increasing with body size); cardiac region slightly elevated, without transverse carina.
Chelipeds ( Figs 6 View FIGURE 6 A, B; 7B) proportionally smaller than those of males. Merus similar to that of male. Carpus with row of minute denticles on outer distal margin. Palm subquadrate, slightly longer than wide; dorsal margin not carinate, but with 2 or 3 rows of granules, glabrous on dorsal and outer surfaces; outer surface with sparse granules and granular ridge extending onto fixed finger adjacent to ventral margin; ventral surface rounded. Fixed finger not deflexed, slightly curved distally, cutting edge forming flange bearing minute tooth distal to midlength and small subterminal concavity to receive tip of dactylus. Dactylus slightly curved, not deflexed, terminating in acute tip crossing with tip of fixed finger; dorsal margin granular; cutting edge with small subtriangular tooth at about midlength.
Ambulatory legs ( Fig. 6 View FIGURE 6 A, B) slightly shorter proportionally, less setose than in male. Fourth pereopod with merus 2.1–2.4 times longer than wide; propodus 1.0–1.2 times wider than long; dactylus about 3.7 times longer than wide. Fifth pereopod with merus 2.4 times longer than wide.
Thoracic sternum conspicuously wide; genital openings small, each with rounded operculum.
Female abdomen ( Fig. 6 View FIGURE 6 B) of 6 free somites, wide, subovate, completely covering thoracic sternum. Somite 1 narrowest, somite 4 widest; somites 3–6 subequal in length. Telson ( Fig. 7 View FIGURE 7 C) transversely trapezoidal, about 4.0 times wider than long, terminal margin truncate.
Pre-adult female (nontype). Carapace about 1.7 times wider than long; granules on branchial ridges conspicuous on lateral sides. Second to fourth pereopods more slender, less setose than in adult females, granules on surfaces sharper; fourth pereopod merus 2.9 times longer than wide, propodus 1.2 times longer than wide; fifth pereopod stout similar to adult females. Abdomen relatively narrow, partially covering thoracic sternum, showing pre-adult condition; telson subsemicircular, about twice wider than long.
Coloration in life. Carapace and pereopods mottled with dirty yellowish brown dorsally, pereopods pale gray ventrally; setae on carapace, chelipeds and ambulatory legs dirty brown; thoracic sternum and abdomen white (see Figs 2 View FIGURE 2 , 6 View FIGURE 6 ).
Distribution and habitat. So far only known from the Obitsu River ( Banzu ) estuary, Kisarazu, Chiba Prefecture; lower intertidal. Found to inhabit the U-shaped tubes of Chaetopterus cautus . The holotype male (CBM-ZC 12293) and paratype female (CBM-ZC 12294) co-inhabited in the same tube of the worm, whereas the female paratype (CBM-ZC 12295) and the pre-adult female (CBM-ZC 12296) were solitary in other tubes of C. cautus .
Remarks. Pinnixa banzu n. sp. appears closest to P. occidentalis from the west coast of North America (Alaska to Baja California). Shared characters include a branchial ridge delimiting anterolateral margin of carapace extending from posterolateral to orbit to branchial lateral region of carapace; a cardiac ridge present at least in the male; a male chela with strongly oblique cutting edge of fixed finger bearing a conspicuous tooth at its midlength and strongly arcuate dactylus; fifth pereopod when extended overreaching distal end of merus of fourth pereopod; male abdomen not conspicuously slender, narrowing distally to roundly triangular or subsemicircular telson that consists of six free somites; male first gonopod with distally flared terminal process. The following characters are useful to differentiate between the new species and P. occidentalis (cf. Rathbun 1918; Zmarzly 1992): (1) the cardiac ridge is completely absent in the female in P. banzu n. sp., whereas there is a still clearly delimited cardiac ridge on the carapace of P. occidentalis , even in the female; (2) the flexor margins of the meri of the second to fourth pereopods are nearly smooth or scarcely granular or spinulose in P. banzu n. sp., but the margins are strongly granular or spinulose in P. occidentalis ; (3) the terminal process of the male first gonopod is stout and bears a distinct shoulder near the base of the chitinous portion in P. banzu n. sp., whereas it is slender and slightly constricted in P. occidentalis ; there is no shoulder on the outer margin near the base of the chitinous part (cf. Zmarzly 1992: Fig. 12E). The habitat also appears to be different in the two species. All specimens of P. banzu n. sp. were found in tubes of the polychaete Chaetopterus cautus , but P. occidentalis has been reported from burrows of echiuran worms or to apparently living freely ( Hart 1982).
Pinnixa chaetopterana View in CoL , known from the western Atlantic (Massachusetts, U.S.A. to Brazil), is also substantially similar to P. banzu View in CoL n. sp. in the most characters cited above. The former is also known to be associated with the chaetopterid worm, Chaetopterus variopedatus (Reiner, 1804) View in CoL . Nevertheless, the new species differs from P. chaetopterna in the following characters (cf. Rathbun 1918; Williams 1984; Martins & d’Incao 1996): (1) a granular ridge extends from the area posterior to the orbit to the lateral part of the carapace in P. banzu View in CoL n. sp., whereas such a ridge is not present in P. chaetopterana View in CoL ; (2) regions of the carapace is not clearly defined in P. banzu View in CoL n. sp., rather than clearly defined by pubescent grooves in P. chaetopterana View in CoL ; (3) the transverse ridge on the carapace is low in P. banzu View in CoL n. sp. even in the male, whereas it forms two dentiform prominences in P. chaetopterana View in CoL ; (4) the subbranchial regions of the carapace is less advanced in P. banzu View in CoL n. sp. than in P. chaetopterana View in CoL ( Figs 3 View FIGURE 3 A, 7A versus Williams 1984: Fig. 360a); (5) the flexor margins of ischium and merus of the fourth pereopod (third ambulatory leg) is sparsely granulate, but not dentate in P. banzu View in CoL n. sp., rather than distinctly dentate in P. chaetopterana View in CoL ; (6) the male abdomen is relatively wider in P. banzu View in CoL n. sp. than in P. chaetopterana View in CoL ( Fig. 3 View FIGURE 3 F. versus Rathbun 1918: Fig. 94b).
Among the congeners known from East Asia, Pinnixa rathbuni View in CoL is most similar to P. banzu View in CoL n. sp., in particular in the general shape of the carapace ( Figs 3 View FIGURE 3 A, 7A and Fig. 8 View FIGURE 8 A) and the structure of the chela ( Figs 4 View FIGURE 4 C, 7B and Fig. 8 View FIGURE 8 B, D) and of the male abdomen ( Fig. 3 View FIGURE 3 F and Fig. 8 View FIGURE 8 C). In fact, P. rathbuni View in CoL had been confused with P. occidentalis View in CoL prior to Sakai (1934) ( Yokoya 1928, 1933). Nevertheless, Pinnixa banzu View in CoL n. sp. can be readily distinguished from P. rathbuni View in CoL by several morphological characters: (1) the carapace is provided with a sharp, bi-sectioned cardiac ridge in the male in P. banzu View in CoL n. sp. ( Fig. 3 View FIGURE 3 A), which is absent in P. rathbuni View in CoL ( Fig. 8 View FIGURE 8 A); (2) the anterolateral to lateral parts of the carapace bear dense short setae in P. banzu View in CoL n. sp. ( Figs 2 View FIGURE 2 A, C, 3A, 6A), rather than being almost glabrous in P. rathbuni View in CoL ( Fig. 8 View FIGURE 8 A); (3) the subbranchial regions of the carapace are more advanced in P. banzu View in CoL n. sp. than in P. rathbuni View in CoL ( Fig. 3 View FIGURE 3 A versus Fig. 8 View FIGURE 8 A); (4) the ambulatory legs are much stouter in P. banzu View in CoL n. sp. than in P. rathbuni View in CoL ( Fig. 4 View FIGURE 4 D, E, G, I versus Fig. 9 View FIGURE 9 A–C, E); for example, length/width of the merus of the fourth pereopod is 2.1–2.4 in P. banzu View in CoL n. sp. (3.2–3.5 in P. rathbuni View in CoL ); (5) setae on the meri to propodi of the second to fifth pereopods are much denser in P. banzu View in CoL n. sp. ( Fig. 2 View FIGURE 2 A–C) than in P. rathbuni View in CoL ( Sakai 1934: text-fig. 1; 1976: text-fig. 323c); (6) flexor carinae on the meri of the third and fourth pereopods are only weakly granular in P. banzu View in CoL n. sp. ( Fig. 4 View FIGURE 4 E, G), rather than noticeably granular or spinulose in P. rathbuni View in CoL ( Fig. 9 View FIGURE 9 B, C); (7) the male abdomen is relatively wider in P. banzu View in CoL n. sp. than in P. rathbuni View in CoL ( Fig. 3 View FIGURE 3 F versus Fig. 8 View FIGURE 8 C; e.g., somite 4 is 3.3 times as wide as long (2.7 times in P. rathbuni View in CoL ); (8) the female telson is terminally truncate in P. banzu View in CoL n. sp. ( Fig. 7 View FIGURE 7 C), rather than broadly rounded in P. rathbuni View in CoL ( Fig. 8 View FIGURE 8 E); (9) the male first gonopod is less strongly tapering distally in P. banzu View in CoL n. sp. than in P. rathbuni View in CoL ( Fig. 5A View FIGURE 5. A – D versus Fig. 5 View FIGURE 5. A – D E); (10) the terminal process of the male first gonopod is more stout and more strongly bent in P. banzu View in CoL than in P. rathbuni View in CoL ( Fig. 5B View FIGURE 5. A – D versus Fig. 5 View FIGURE 5. A – D F). Furthermore, the new species may attain maturity in the female at the greater size than in P. rathbuni View in CoL . The female specimens of P. rathbuni View in CoL examined in this study (3.7 × 7.2 to 4.4 × 7.8 mm), equivalent in the size to the smallest premature female specimen of P. banzu View in CoL n. sp. (4.1 × 7.1 mm), are all fully mature.
Pinnixa rathbuni View in CoL has been reported as a free-living species on soft sediments ( Sakai 1976; Sekiguchi 1983), although studies in the northern part of the Sea of Japan and the Sea of Okhotsk ( Kobjakova 1967; Vassilenko 1990; Kornienko & Korn 2010) recorded an association with Chaetopterus View in CoL species (see below). The comparative specimens of P. rathbuni View in CoL reported herein were all collected by SM grab from soft sediments, and no association with other invertebrates was recorded.
The small pre-adult female of P. banzu View in CoL n. sp. (CBM-ZC 12296) differs from adult females (CBM-ZC 12294, 12295) in the proportionally narrower carapace, relatively slender ambulatory legs, and the narrower abdomen only partially covering the thoracic sternum. The ontogenetic change in the carapace shape and the stoutness of the ambulatory legs is known for other congeneric species ( Rathbun 1918; Williams 1984; Zmarzly 1992), although these characters are diagnostic for adult specimens. These characters influenced by ontogenetic change should be used with caution when identifying juvenile or subadult specimens.
Kobjakova (1967), Vassilenko (1990) and Kornienko & Korn (2010) reported the commensal association of P. rathbuni View in CoL with Chaetopterus View in CoL species from the continental margins of the Sea of Japan and the Sea of Okhotsk. Considering the superficial resemblance between P. banzu View in CoL n. sp. and P. rathbuni View in CoL and the association of the new species with Chaetopterus cautus View in CoL , it is likely that these Russian records might be referred to the new species. Kornienko & Korn (2010) discussed larval differences between their material and the previous descriptions of P. rathbuni View in CoL by Japanese workers ( Sekiguchi 1978; Muraoka 1979; Konishi 1983). The photo of an adult crab given by Kornienko & Korn (2010: 179) actually agrees with the new species, rather than with P. rathbuni View in CoL , in particular in the proportion of the ambulatory legs and the color in life. Examination of voucher specimen(s) is needed to satisfactorily evaluate the specific identity of these Russian records.
The following nine species presently assigned to Pinnixa View in CoL (except for P. rathbuni View in CoL ) are known to be associated with species of Chaetopterus View in CoL : P. bahamondei Garth, 1957 View in CoL , P. brevipollex Rathbun, 1898 View in CoL , P. chaetopterana Stimpson, 1860 View in CoL , P. da r w i ni Garth, 1960, P. sayana Stimpson, 1860 View in CoL , P. tomentosa Lockington, 1877 View in CoL , P. transversalis (H. Milne Edwards & Lucas, 1844) View in CoL , P. t ub i c o l a Holmes, 1894, P. valdiviensis Rathbun, 1907 View in CoL ( Schmitt et al. 1973; Zmarzly 1992; Coelho 1997; Petersen & Britayev 1997). The present new species represents the tenth example of the genus associated with Chaetopterus View in CoL tube worm.
Etymology. Named after the type locality, Banzu , Kisarazu, Chiba Prefecture, Japan. Used as a noun in apposition.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pinnixa banzu
| Komai, Tomoyuki, Nishi, Eijiroh & Taru, Masanori 2014 |
P. bahamondei
| Garth 1957 |
P. valdiviensis
| Rathbun 1907 |
P. brevipollex
| Rathbun 1898 |
P. tomentosa
| Lockington 1877 |
P. chaetopterana
| Stimpson 1860 |
P. sayana
| Stimpson 1860 |
P. transversalis
| H. Milne Edwards & Lucas 1844 |
Chaetopterus variopedatus
| Reiner 1804 |