
Phyxioschema erawan, Schwendinger, Peter J., 2009
|
publication ID |
https://doi.org/ 10.5281/zenodo.188258 |
|
DOI |
https://doi.org/10.5281/zenodo.6214226 |
|
persistent identifier |
https://treatment.plazi.org/id/03A9B15A-FFA0-7E7A-FF06-B4E2FF2DF946 |
|
treatment provided by |
Plazi |
|
scientific name |
Phyxioschema erawan |
| status |
sp. nov. |
Phyxioschema erawan sp. n.
Figures 4–7 View FIGURE 4 A – E View FIGURE 5 A – E View FIGURE 6 A – L View FIGURE 7 A – H , 22A View FIGURE 22 A – I
Phyxioschema suthepium , group B: Raven & Schwendinger 1989: 56, 59 (misidentification).
Material: THAILAND (western central region): Kanchanaburi Province, Si Sawat District, Erawan National Park, below Phrathat Cave (14°23’47”N, 99°05’05”E), 560–650 m, male holotype (matured 22.XII.2006), 5 male paratypes (matured 20.XII., 24.XII.2006, 16.I., 20.I., 9.II.2007), 7 female paratypes, 1 juvenile male, collected by P.J. Schwendinger, 12.XII.2006 (sample TH-06/12), MHNG; same locality (14°23.909’N, 99°04.893’E), 590 m, 1 female paratype, collected by L. Monod, 7.IX.2001 (sample TM-01), MHNG; same locality (14°23’51”N, 99°04’55”E), 600 m, 1 male paratype (decayed and fragmented; matured in January 2006; n° O-28B; SMF), 2 female paratypes (n° O-28A, SMF; n° O-28C, "allotype", MHNG), collected by S. Huber on 16.XI.2004.
THAILAND (northern region): Chiang Mai Province, along road from Chiang Dao to Chai Prakan, at border between both districts (19°35'32"N, 99°07'11"E), 500–770 m, 9 male paratypes (two collected mature, others matured on 20.XII., 31.XII.2007, 21.I., 4.II., 13.II.2008, late XII.2008, 15.I.2009), 22 female paratypes, 19.+ 21.XII.2007 (sample TH-07/16); same locality but given as "Fang, 750 m " in Raven & Schwendinger (1995: 56, 59), 1 male paratype (matured on 19.XII.1989, moulted again (!) and died on 5 May 1990) collected 31 October 1987 and 1 female paratype collected 27 August 1990. All specimens from this locality collected by P.J. Schwendinger and deposited in MHNG.
Etymology: The specific epithet, a noun in apposition, refers to the Erawan National Park, named after the three-headed Elephant of the Thai mythology, which corresponds to the mythical elephant Airavata in the Hindu religion.
Diagnosis: This is a medium-sized species with short, stout legs and metatarsal preening combs. It is distinguished from all other known Phyxioschema species by males having only tarsi III and IV pseudosegmented (all leg tarsi either integral or pseudosegmented in other species). Males differ from those of the smaller P. suthepium by a relatively longer and more slender palpal tibia, by the absence of a triangular projection on the retrolateral margin of patella I, by the presence of retroventral and retrolateral spines on tibia I, by a longer band of hooked spinules retrodorsally on femur I, by a more prominent retroventral keel on metatarsus II, and by a prolaterally slightly more incrassate tibia II with a less curved and less inclined band of elongated prolateral spinules. Females differ from those of P. s u t h e p i u m by median receptacles composed of shorter, straight or only moderately convoluted median stalks and heads without pores, and by lateral receptacles basally constricted to a neck; secondary receptacles are usually absent.
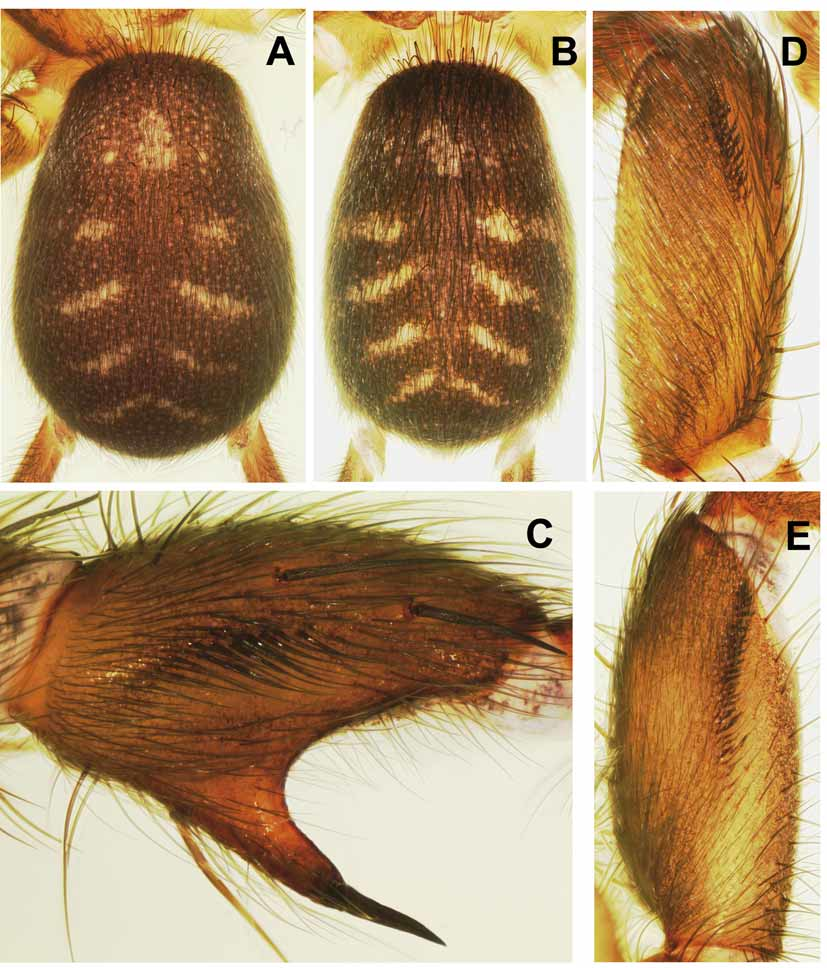
Description: MALE (holotype). Colour in alcohol: Carapace brown, mottled with dark brown, eye mound black. Sternum, chelicerae, leg coxae and trochanters (dorsally and ventrally) light brown, remaining leg articles contrasting, dark brown; palps cream. Labium and maxillae light brown, with white anterior and prolateral zones, respectively. Opisthosoma blackish brown, with an indistinct pattern of light patches and light transverse stripes on dorsal side ( Fig. 4B View FIGURE 4 A – E ); genital region and booklung plates light brown; spinnerets blackish brown, with light spots ventrally.
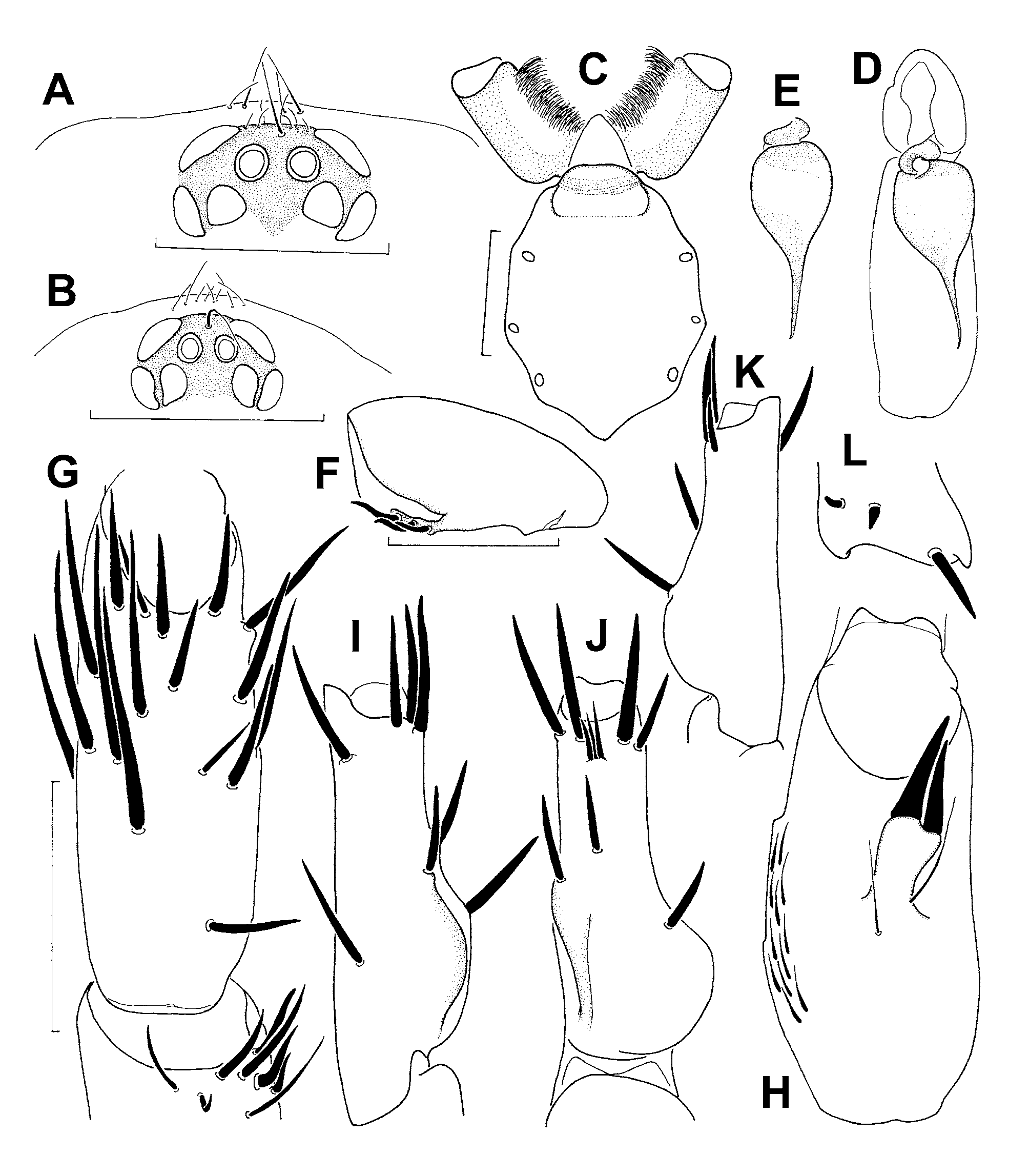
Body 9.0 long. Carapace 4.7 long, 4.2 wide, oval, almost flat, covered with fine dark adpressed hairs (most straight, few slightly curved), these densest behind eye mound; few stronger bristles on and in front of eye mound and on posterior margin. Eyes on low mound; eye group 0.37 long, anterior eye row slightly procurved, 0.59 wide, posterior eye row very slightly recurved, 0.62 wide ( Fig. 6B View FIGURE 6 A – L ). Eye diameters and interdistances: AME 0.13, ALE 0.27, PME 0.18, PLE 0.19; AME–AME 0.04, AME–ALE 0.02, PME–PME 0.18, PME–PLE 0.02. MOQ 0.30 long, 0.27 wide anteriorly, 0.41 posteriorly. Fovea pitlike, with 2 long, thickened and slightly procurved foveal setae anterior to it.
Chelicerae weak, grooves with 9/10 prolateral teeth and 15/16 medioproximal denticles. Maxillae ( Fig. 6C View FIGURE 6 A – L ) 0.8 long, 0.5 wide; with pallid prolateral zone; anterior lobe indistinct, with fairly wide but indistinct serrula on ridge. Labium ( Fig. 6C View FIGURE 6 A – L ) 0.3 long, 0.6 wide, anterior edge distinctly setose, followed by pallid zone; posterior part pigmented, with few fine setae. Sternum ( Fig. 6C View FIGURE 6 A – L ) 1.8 long, 1.5 wide, cordate, with deeply excavated post-labial depression formed by fused anterior sigilla and labio-sternal groove, and with 3 pairs of indistinct marginal sigilla. Membrane at posterior sternal exfoliations (at height of coxae III and IV) with dark pigment.
Palps ( Fig. 6D–E View FIGURE 6 A – L ) set with long stiff bristles. 7+7 trichobothria in 2 rows on tibiae, about 10 in a zig-zag row on tarsi. Palpal bulb with oval base and quite long, tapering, almost straight embolus with slightly curved tip.
Legs 2134. All tarsi armed with spines. Tarsi I–II integral, III–IV pseudosegmented. Distoventral preening combs (composed of 2–3 setae) on metatarsi I–IV (on IV retroventrally). I: tibia cylindrical, with strong spines prolaterally, ventrally and retrolaterally ( Fig. 6G View FIGURE 6 A – L ); patella with row of 3 short sigmoid spines retroventrally (followed by 2 longer curved spines on distoventral margin), without triangular projection on retrolateral margin ( Fig. 6F View FIGURE 6 A – L ); femur I with relatively long and narrow band of hooked spinules retrodorsally ( Fig. 4D View FIGURE 4 A – E ; better visible in paratype, Fig. 5D View FIGURE 5 A – E ). II: metatarsus ventroproximally with 2 widely separated, rounded and bulged (not sharp) keels ( Fig. 6I –L View FIGURE 6 A – L ); the proventral one ( Fig. 6I, K View FIGURE 6 A – L ) shorter and ventrally directed; the other one ( Fig. 6J–L View FIGURE 6 A – L ) longer and retroventrally directed; tibia distinctly incrassate, band of elongated spinules on prolateral side moderately curved, slightly inclined from longitudinal axis of tibia, fairly long, reaching beyond height of distal side of ventral spur ( Fig. 4C View FIGURE 4 A – E , see also Fig. 5C View FIGURE 5 A – E ); ventral spur of tibia with widened and slightly bilobed apex carrying 2 megaspines, these almost parallel to each other, prolateral megaspine distinctly surpassing retrolateral one ( Fig. 6H View FIGURE 6 A – L ); femur with a long and narrow band of hooked spinules proventrally ( Fig. 4E View FIGURE 4 A – E ; better visible in paratype, Fig. 5E View FIGURE 5 A – E ). Spination: I: patella v2 /3, r3; tibia p3, v9/10, r2/4; metatarsus v9; tarsus p2, v0/1, r 3. II: patella p2; tibia p2, v1 +2 megaspines; metatarsus d2, p2, v5; tarsus p2, v0/1, r 3. III: patella p3, r1/2; tibia d2, p2, r2, v6 /7; metatarsus d4/5, p2/3, r1, v8; tarsus p2, r 2. IV: patella p2/3, r1; tibia d2, p2, r2, v6 /7; metatarsus d6, p3, r1, v8; tarsus p2, r2. Trichobothria: 8+ 8 in 2 rows on tibiae; ca 10 (difficult to see) in one row on metatarsi; ca 10 (difficult to see) in one row on tarsi. Paired claws with 8–10 teeth in sigmoid row, unpaired claw with 4–6 sessile teeth.
Opisthosoma 4.2 long, 2.7 wide; dorsal side quite densely covered with many fine adpressed black hairs and fewer stiff black bristles with darkened sockets ( Fig. 4B View FIGURE 4 A – E ); ventral side only with fine black hairs. Posterior median spinnerets 0.5 long; posterior lateral 4.5 long (proximal article 1.2, median article 1.4, FEMALE ("allotype"). As the male, except for: whole body more strongly bleached (probably due to longer preservation in alcohol); prosoma (especially chelicerae, maxillae and palpal tarsi) darker; opisthosoma with more distinct dorsal pattern of four pairs of light transversal stripes forming incomplete chevrons in posterior two thirds, and of one pair of larger light patches in anterior third ( Fig. 4A View FIGURE 4 A – E ); pale prolateral zone of maxillae and pale anterior zone of labium more distinct.
Habitus, see Fig. 22A View FIGURE 22 A – I . Body 12.5 long. Carapace 4.3 long, 3.4 wide. Eye group ( Fig. 6A View FIGURE 6 A – L ) 0.49 long, anterior eye row 0.80 wide, posterior eye row 0.86 wide. Eye diameters and interdistances: AME 0.15, ALE 0.33, PME 0.19, PLE 0.27; AME–AME 0.06, AME–ALE 0.03, PME–PME 0.24, PME–PLE 0.02. MOQ 0.36 long, 0.36 wide anteriorly, 0.58 posteriorly.
Chelicerae stronger than in male, grooves with 10/11 prolateral teeth and 27/28 median proximal denticles. Maxillae 1.3 long, 0.8 wide, serrula slightly wider than in male. Labium 0.4 long, 0.8 wide. Sternum 2.0 long, 1.8 wide.
Palps with 7+7 trichobothria on tibiae and 11 on tarsi. Tarsal claw with 12 teeth.
Legs 2314. All tarsi integral. Spination: I: patella p1; tibia p1, v4; metatarsus v8; tarsus p2, r 2. II: patella p2; tibia p2, v5; metatarsus p2, v8 /9; tarsus p2, r 2. III: patella p3/5, r2; tibia d2, p2/3, r2, v4; metatarsus d4, p3, r2, v9; tarsus p2, r 2. IV: patella p2, r1; tibia d2, p2, r2, v4; metatarsus d4, p3, r2, v8 /9; tarsus p2, r2. Trichobothria: 2 rows of 8–9 each on tibiae; 12 in one on metatarsi; 9 in one row on tarsi. Paired claws with 9–11 teeth in sigmoid row, unpaired claw with 3–4 sessile teeth.
Opisthosoma 6.7 long, 4.4 wide. Posterior median spinnerets 0.6 long; posterior lateral spinnerets 5.4 long (proximal article 1.6, median article 1.6, distal article 2.2).
Vulva ( Fig. 7C View FIGURE 7 A – H ) with moderately wide spermathecae, each with 2 receptacles; base of lateral receptacle slightly constricted to neck, with sclerotisation only on ental side, head of lateral receptacle densely covered with pores; median receptacle with a short, slightly curved, completely sclerotised stalk and a globular head without pores; secondary receptacles absent.
Palp and leg measurements: See Table 1.
Variation: Measurements of males (n=17), of females with egg sacs (n=5) and of largest female (in parentheses): body length 6.9–8.3, 7.3–12.3 (13.3), carapace length 2.9–3.4, 3.0–4.3 (5.0), width 2.3–2.7, 2.4–3.4 (4.0). Specimens from the northern population usually possess a more distinct dorsal opisthosomal pattern. This is composed of four incomplete light chevrons in the posterior two-thirds, plus an unpaired (usually paired in the southern population; Fig. 4A View FIGURE 4 A – E ), fairly large median patch, often laterally flanked by a pair of smaller patches, situated in the anterior third ( Fig. 5A–B View FIGURE 5 A – E ). Specimens from the southern population are generally larger than those from the northern population: body length, carapace length and carapace width in males is 7.7–8.3, 3.2–3.4, 2.5–2.7 (n=7; south) versus 6.9–8.5, 2.9–3.2, 2.3–2.6 (n=10; north). One male has only a single foveal seta, three females have three foveal setae, all other specimens examined have two. Tarsus III of males is more or less distinctly pseudosegmented. One female has no preening combs on metatarsus I; one male lacks preening combs and proximoventral spines on metatarsus II of one side. One male has only two sigmoid spines (most have three) retroventrally on patella I of one side; four males have four such spines in a row, mostly on one side (one male on both sides); one male has a fourth such spine situated ventrally to the usual row of three spines on one side. Female genitalia are quite variable. No variation was observed in the number of megaspines on the ventral spur of tibia II; all 17 males examined possess two megaspines on both sides. In females from the southern population ( Fig. 7A–D View FIGURE 7 A – H ), the median receptacles usually have shorter stalks than in females from the northern population ( Fig. 7E–H View FIGURE 7 A – H ). Three females have three receptacles on one side. In one of them, the third receptacle arises from the base of the median receptacle ( Fig. 7H View FIGURE 7 A – H ) and thus is a duplication rather than a separate secondary receptacle; in the second female this also appears to be the case ( Fig. 7D View FIGURE 7 A – H ). The third female has a small duplication of a lateral receptacle ( Fig. 7G View FIGURE 7 A – H ), which is clearly abnormal and was never observed in any other Phyxioschema females.
Relationships and taxonomic status: Phyxioschema erawan sp. n. appears most closely related to P. suthepium . Both species share the presence of metatarsal preening combs (absent in other congeners), a dark colouration, a fairly distinct dorsal opisthosomal pattern, relatively short stout legs, a distinctly incrassate tibia II and very similar modifications of leg II in males; moreover, both species live on the ground.
Remarks: The presence of two distinct species in northern Thailand was reported by Schwendinger (1988: 234). In Raven & Schwendinger (1989), however, specimens of the northern population (between Chiang Dao and Chai Prakan) of P. erawan sp. n. were regarded only as a variant form (group B) of P. suthepium on the basis of different behaviour. Morphological characters for a distinction of both species were not apparent because female genitalia were not thoroughly studied.
Distribution and habitat: This species was found at two localities separated by about 580 km ( Fig. 1 View FIGURE 1 , localities 1 and 14); both lie on or close to limestone hills. Phyxioschema suthepium occurs between these localities, and was also found at one locality on limestone [specimens collected on a small, isolated limestone hill at Ban On Luai ( Fig. 1 View FIGURE 1 , locality 3)]. It appears at present that both P. erawan sp. n. populations are widely disjunct and that the species is restricted to extensive limestone areas. At the southern locality ( Erawan N.P.; type locality) webs were found under stones and inside old, broken bamboo poles lying on the floor of a dry deciduous forest, as well as at the base of limestone rocks jutting out of the soil. In the latter case, the silken retreats were built deep into cracks between soil and rock, which made it quite difficult to extract the spiders without damaging them. These webs were particularly visible along the footpath between the park ranger station and the Phrathat Cave, but none were seen on roadsides and earth banks outside the forest. At the northern locality (between Chiang Dao and Chai Prakan) webs were built mostly on roadsides and cut paths in cultivated land close to limestone hills, and all retreats were leading into holes in the soil.
Phenology and reproduction: Males matured between late December and mid-February. The first egg sacs were built about 1.5 months later, usually followed by another four egg sacs per female in intervals of about three weeks until early May; they contained 26– 58 eggs (n = 15). In captivity, some females from the northern population built their egg sacs into depressions on the ground and covered them with a layer of interwoven soil particles. This effective camouflage was employed when the first egg sac (suspended in the web, as usual in Phyxioschema ) was removed, and I did not observe this behaviour in females of the southern population or of other Phyxioschema species. Spiderlings hatched about four weeks after oviposition. A male that hatched from one of the egg sacs in February became mature in mid-December of the same year. Mature females moulted at least twice per year (some up to four times), mostly in January to May and in August to December; reproducing females moulted in March to July and again in September and November. There appears to be only one mating period per year; observations in situ from spring and summer, however, are lacking. See Table 3.
Mating behaviour: Six copulations were observed in captivity; they lasted 16, 23, 25, 34, 37 and 45 min, respectively. During interlocking, both partners held their bodies mostly straight, the male having its opisthosoma slightly bent ventrally (not dorsally as observed in P. s u t h e p i u m) by not more than 30°.
Commensals: Pygmephorid mites, probably belonging to the genus Bakerdania Sasa or to an undescribed genus (S. Mahunka, pers. commun.), were found in the fovea of several spiders (mostly females, including one collected by S. Huber in 2004) from both populations. The thin, strongly elongated, only slightly curved and asymmetrical claws on the posterior legs of these mites indicate that they are adapted to move between the hairs of their hosts. It is quite unlikely that these are soil-dwelling mites that accidentally crawled onto the spiders. No perforations of the spider cuticle in that area (an unsuitable place for that purpose) are discernible; therefore these mites (very small and weakly sclerotised) are obviously not parasites. A non-parasitic association of the pygmephorid mite Pseudopygmephorus atypoides Rack with the burrowing mygalomorph spider Atypoides riversi O.-P. Cambridge ( Antrodiaetidae ) in California was reported by Vincent & Rack (1982).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Phyxioschema erawan
| Schwendinger, Peter J. 2009 |
Phyxioschema suthepium
| Raven 1989: 56 |