
Photoplagios antongil, SPARKS, 2006
|
publication ID |
https://doi.org/ 10.1206/0003-0082(2006)3526[1:ANSOPT]2.0.CO;2 |
|
DOI |
https://doi.org/10.5281/zenodo.5061701 |
|
persistent identifier |
https://treatment.plazi.org/id/03C85256-FF8E-FF8D-1BA6-FC05FE0606FF |
|
treatment provided by |
Felipe |
|
scientific name |
Photoplagios antongil |
| status |
sp. nov. |
Photoplagios antongil View in CoL , new species
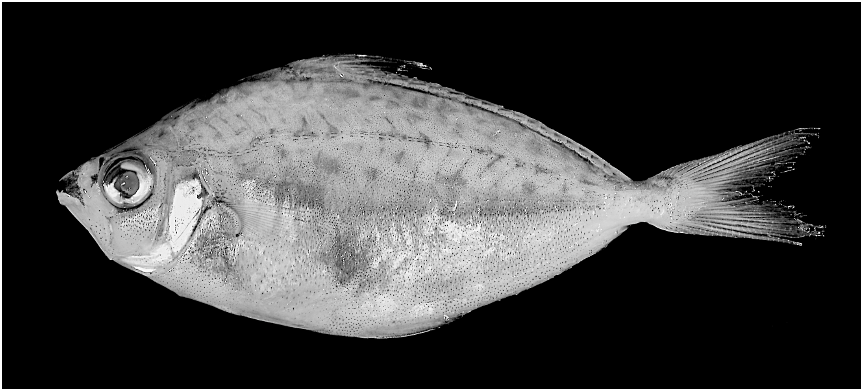
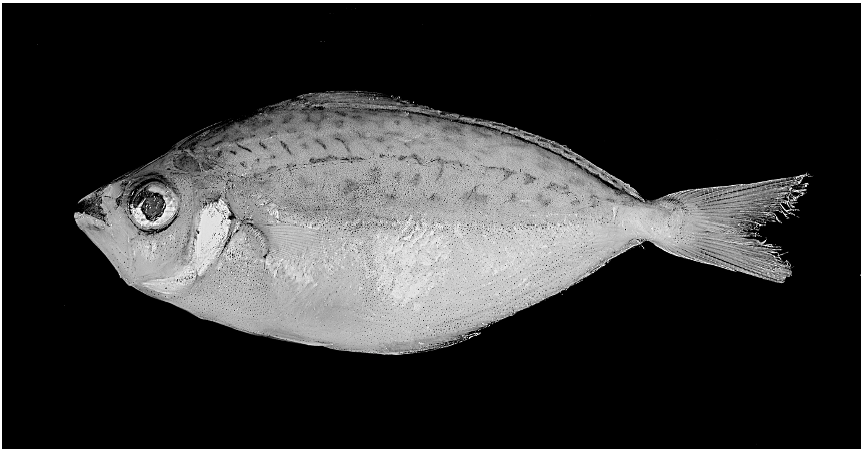
Figures 3 View Fig , 4 View Fig
Photoplagios n. sp. ‘‘Madagascar’’: Sparks et al. (2005).
HOLOTYPE: AMNH 236544 About AMNH , 75.0 mm SL, adult male; Northeastern Madagascar: Antongil Bay : Maroansetra market; J.S. Sparks, W.L. Smith, and K.L. Tang, Mad JSS 29-2003, Nov. 2003.
PARATYPES: AMNH 236545 About AMNH , 3 ex., 77.3– 81.8 mm SL; data as for holotype . SIO 05-117 About SIO , 1 ex., 76.0 mm SL; data as for holotype .
DIAGNOSIS: Males of Photoplagios antongil are distinguished from congeners by the presence of a broad and presumably translucent (in life) midlateral stripe, which is darkly pigmented in preservative due to a concentration of melanophores, and by the presence of two darkly pigmented flank patches (presumably also translucent in life) located ventral to the lateral midline, one anteroventral to the pectoral-fin base and another ventral to the lateral midline at about midbody. Photoplagios antongil is further distinguished from P. leuciscus , the only externally similar species occurring in the region, by the absence of a large translucent triangular patch on the flanks (present in male P. leuciscus ); upper flank pigmentation consisting of large, sparse spots and blotches (vs. highly speckled with fine lines and small spots); a much shorter, although elongate, second dorsal-fin spine; a straight predorsal profile (vs. weakly S-shaped); absence of black pigment in the pectoral-fin axil; and exposed conical oral dentition in two distinct rows (vs. multiple closely set and indistinct rows of unexposed villiform teeth).
DESCRIPTION: Selected proportional measurements and meristic data presented in table 2. A moderately shallow-bodied and elongate leiognathid. Body laterally compressed. Lateral snout outline mostly straight. Weak preorbital protuberance due to protrusion of both frontal and lateral ethmoid ossifications. Predorsal head profile mostly straight to mildly convex. Nuchal spine slightly protruding and distal tip exposed. Nuchal spine with distinct median keel. Two short and stout postnasal spines (5 protuberances) present on lateral ethmoid, located posterior to nasal foramina and just rostrodorsal of orbit. Postnasal spines followed posteriorly by well-developed supraorbital ridges that converge posteriorly. Dorsal and ventral body profiles moderately rounded. Dorsal-fin origin located slightly posterior to vertical through pelvic-fin origin. Anal-fin
TABLE 2 Morphometric and Meristic Data for Photoplagios antongil , New Species
origin located at about level of vertical through the last (5 8th) dorsal-fin spine or first dorsal-fin ray. Eye large. Caudal peduncle slender and shallow. Mouth small and terminal in position, directed slightly downward when protruded. Posterior margin of maxilla exposed, reaching to level of vertical through anterior margin of orbit. Anterior nasal pore small and round; posterior foramen much larger and crescent-shaped, partially encircling anterior pore. Preopercular margin weakly serrate along ventral and ventrocaudal margins. Vertebral count: 10 precaudal + 13 caudal 5 23. Neural and hemal spines of vertebral centrum PU4 somewhat expanded and bladelike. Twelve or 13 elongate and triangular outer gill rakers arrayed along lower limb (5 ceratobranchial one) of first gill arch.
Fins: Dorsal fin with VIII spines and 16 branched rays. First dorsal-fin spine greatly reduced in length and relatively robust. Second through fourth dorsal-fin spines elongate and robust; second spine longest. Second dorsal-fin spine moderately, but not exceedingly, elongate. Third and fourth dorsal-fin spines serrate along anterior margin and ‘‘lock’’ into groove on posterior margin of preceding spine when erect. Dorsal-fin spines five through eight feeble, shorter than second through fourth spines. Anal fin with III spines and 14 branched rays. First anal-fin spine very short. Second and third anal-fin spines robust and elongate; second spine longest, but not appreciably longer than third. Third anal-fin spine serrate on anterior margin and ‘‘locks’’ into groove on posterior margin of second spine when erect. Spinous dorsal and anal fins with asquamate basal sheath. Pelvic fins short, not reaching first anal-fin spine when adducted (i.e., an appreciable gap present). Eight upper and seven lower branched caudal-fin rays. Seventeen total pectoral-fin rays.
Dentition: Two distinct rows of closely set, elongate and recurved conical teeth present in both upper and lower jaws. Upper jaw teeth somewhat larger than those of lower jaw, particularly anteriorly. Lips not fleshy, and teeth exposed.
Squamation: Body scales cycloid, but not remarkably small. Head and opercular region asquamate. Chest fully scaled, except along ventral midline. Scales conspicuous and extend anteriorly ventral to opercle to about anterior margin of chest. Lateral line arched and complete. Pored scales in lateral line number approximately 60 to 63. Scales difficult to count posteriorly due to high proportion missing. Pores well developed. Pelvic axillary scale well developed and elongate. All fins asquamate, except for caudal fin, which bears several rows of scales of reduced size, particularly centrally on fin.
PIGMENTATION IN PRESERVATIVE: Body ground coloration yellowish-olive above lateral midline and creamy pale yellow below. Iridescent silvery patches present to varying degree ventral to lateral midline (although much of guanine lost in preservation). Opercle, subopercle, and interopercle iridescent and silvery. Midlateral stripe present in males. Stripe broad and darker than flank regions above and below due to concentration of melanophores, and presumably translucent in life (compare figs. 3 and 4, illustrating an adult male and female, respectively). Stripe begins posterodorsal to pectoral-fin base and extends to about anterior margin of caudal peduncle. Two dark gray to charcoal, and also presumably translucent in life, patches present on flank in males, one anteroventral to pectoral-fin base in all specimens and another ventral to lateral midline at about midbody in only some specimens (compare figs. 3 and 4). Based on comparisons to the translucent flank patches in congeners (e.g., P. leuciscus , P. klunzingeri , P. stercorarius , and P. moretoniensis ) in various states of preservation, in specimens in which the silvery guanine layer has faded in preservation, these flank patches are generally very obvious due to a concentration of melanophores causing them to appear blackish (see fig. 5). Thus, it is reasonable to conclude that the blackish midlateral stripe and two lower flank patches in the new species, in which the silvery guanine on the integument is mostly lost in preservation in all specimens, are also translucent in life.
Pigmentation pattern above lateral midline characteristically blotchy and mottled. Blotches large, variable in size, and sparsely arranged. Longitudinal series of blackish spots arrayed directly above midlateral stripe. Pores of lateral line scales edged dorsally and ventrally with melanophores; in combination with concentration of blackish blotches present along lateral line, melanophores create an arching and irregular stripe. Body ventral to lateral midline peppered with melanophores. Cheek and gular region pale yellow; some silvery iridescence on cheek. Head above orbit and nape grayish to grayish brown. Snout with large black patch above upper lip. Lips pale yellow. Two distinct black blotches present anterior to orbit; dorsal blotch covering postnasal spines. Line of black pigment ventral to eye. Chest and belly gray, silvery, or pale yellow; silvery along ventral midline. Caudal peduncle silvery with blackish saddle on dorsal margin. Base of caudal fin iridescent and silvery. Dorsal and anal fins hyaline to pale white, with a concentration of black pigment distally on dorsal-fin spines. Pectoral and pelvic fins pale yellow. Black stripe present on body ventral to dorsal-fin base; more prominent posteriorly. Caudal fin yellowish proximal to base, white distally. Membrane of caudal fin with concentration of black pigment, creating series of thin black radiating lines.
LIGHT- ORGAN SYSTEM (LOS): Sexual dimorphism of the light organ and associated internal structures is evident, but not striking as in some other members of the genus (e.g., P. elongatus and P. rivulatus [ Dunlap and McFall-Ngai, 1984; Sparks et al., 2005]). The light organ of males in general is enlarged compared to similarly sized conspecific females. The dorsal lobes of the light organ in particular are enlarged in males, but not greatly, and extend into gas bladder only slightly at the light-organ window (fig. 2). The light organ itself is highly spotted, with prominent black melanophores on a yellow background, lending a leopard skin pattern to much of the light organ. Based on the limited material available, the light organ of females appears in general to be smaller and not as densely spotted, but more solidly black. There is a broad lateral clearing of the gas bladder lining, extending almost the entire length of the gas bladder chamber, and the chamber is silvered laterally only at the anterior end near the light-organ window. The gas bladder is peppered with melanophores anteriorly near the light-organ window and also caudoventrally, but there are no melanophores present in the guanine-free (5 clear) region of the gas bladder lining.
The material available is not ideally preserved for reliably sexing some of the specimens (i.e., the gonads are poorly preserved) or for interpreting the extent of flank pigmentation, translucence, or silvering in life. Therefore, until additional material becomes available, I am reluctant to comment further on external sexual dimorphism of the LOS apart from noting that in specimens of Photoplagios (and Photopectoralis ) in which the silvery guanine is lost in preservative, as it is in the type series of P. antongil , regions that were translucent in life generally appear blackish due to a concentration of dispersed melanophores (fig. 5B and C). It would appear that these concentrated melanophores, which are generally aggregated and quite small in well-preserved specimens in which the guanine layer is intact (fig. 5A), can be rapidly dispersed to occlude the transparent flank patches and stripes so as to inhibit the emission of luminescence from the light organ.
DISTRIBUTION: Known only from market specimens purchased in northeastern Madagascar (Maroansetra market). The Malagasy fishermen who supply the market work locally in Antongil Bay , and it can safely be assumed that is where the type series was collected. Presumably the species is more widely distributed; however, it is not represented in collections made elsewhere in Madagascar that are available to the author. In their guide to the commercial fishes of Madagascar, Bauchot +
market. Specimen with silvery guanine intact. Flank patch difficult to discern unless specimen is examined obliquely. (B) AMS I.22978004, adult male, 95.8 mm SL; northern Australia. Specimen in which guanine is somewhat faded. Flank patch is easy to locate as darker triangular area surrounded by silvery integument. (C) USNM 373280, adult male, 79.8 mm SL; Iran. Specimen in which guanine is completely lost in preservative. Triangular flank patch appears black due to concentration of dispersed melanophores (see text for discussion).
and Bianchi (1984) list Leiognathus lineolatus as present in the region. The fish referred to as L. lineolatus may correspond to the new species; however, no specimens from the study of Bauchot and Bianchi (1984) appear to have been deposited at MNHN.
ETYMOLOGY: Named in reference to the type locality, Antongil Bay , in northeastern Madagascar. The specific epithet, antongil , is used as a noun in apposition .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Order |
|
|
Family |
|
|
Genus |