
Philinopsis falciphallus, Gosliner, 2011
|
publication ID |
https://doi.org/10.11646/zootaxa.2751.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/394D87A1-FFFD-234C-FF5E-FC07F87D6F56 |
|
treatment provided by |
Felipe |
|
scientific name |
Philinopsis falciphallus |
| status |
sp. nov. |
Philinopsis falciphallus View in CoL n. sp.
( Figures 1A, B View FIGURE 1 , 2A View FIGURE 2 , 3 View FIGURE 3 , 4 View FIGURE 4 )
Philinopsis sp. 5 Gosliner et al., 2008: 42, upper photo.
Philinopsis sp. Rosenfeld, 1999 ; Abbott, 2007; Miller, 2008.
Aglajid sp. e518 Johnson, 2008.
Aglajid sp. 7 Rudman, 2005c.
Material examined. Holotype: CASIZ 181228 , 12 m depth, Mainit Bubbles , Mabini, Batangas Province, Luzon, Philippines, 13.686025°S, 120.895167° E, 20 May 2009, T. M. Gosliner GoogleMaps . Paratype: CASIZ, 177678, one specimen, dissected, 3 m depth, Matutunggil Point , Mabini, Batangas Province, Luzon, Philippines, 13.756132°S, 120.906889° E, 19 April 2008 GoogleMaps , T. M. Gosliner . CASIZ 174145 , one specimen, dissected, 12 m depth, Mainit Bubbles , Mabini, Batangas Province, Luzon, Philippines, 13.686025°S, 120.895167° E, 24 May 2009 GoogleMaps , M. D. Miller. CASIZ 180406 , one specimen, 3 m depth, old site of Seafari Resort , also known as Basura , Anilao , Mabini, Batangas Province, Luzon, Philippines, 13.756788°S, 120.921605° E, 17 May 2009 GoogleMaps , T. M. Gosliner . CASIZ 182236 , one specimen, dissected, 8 m depth, on Halimeda, Bigej-Meck Reef , Kwajalein Atoll, Marshall Islands, Scott Johnson , 21 April 2008 . CASIZ 182237 , one specimen, 9 m depth, on Halimeda, Bigej-Meck Reef , Kwajalein Atoll, Marshall Islands, Jeanette Johnson , 16 November 2008 .
Geographical distribution. Marshall Islands ( Johnson 2008), Philippines ( Rosenfeld 1999; Miller 2008; Lee 2008), Indonesia ( Abbott 2007), Red Sea ( Koretz 2005a).
Etymology. The name “falciphallus” is a combination of the Latin falcis, meaning sickle and the Greek phallos, meaning penis. This refers to the sickle-shaped structure associated with the penis of this species.
Natural history. This species is found on coral rubble in 5–15 m depth and has been observed depositing egg masses at the base of Halimeda plants ( Johnson 2008). A specimen from the Red Sea was observed apparently feeding on polyclad flatworms ( Koretz 2005a).
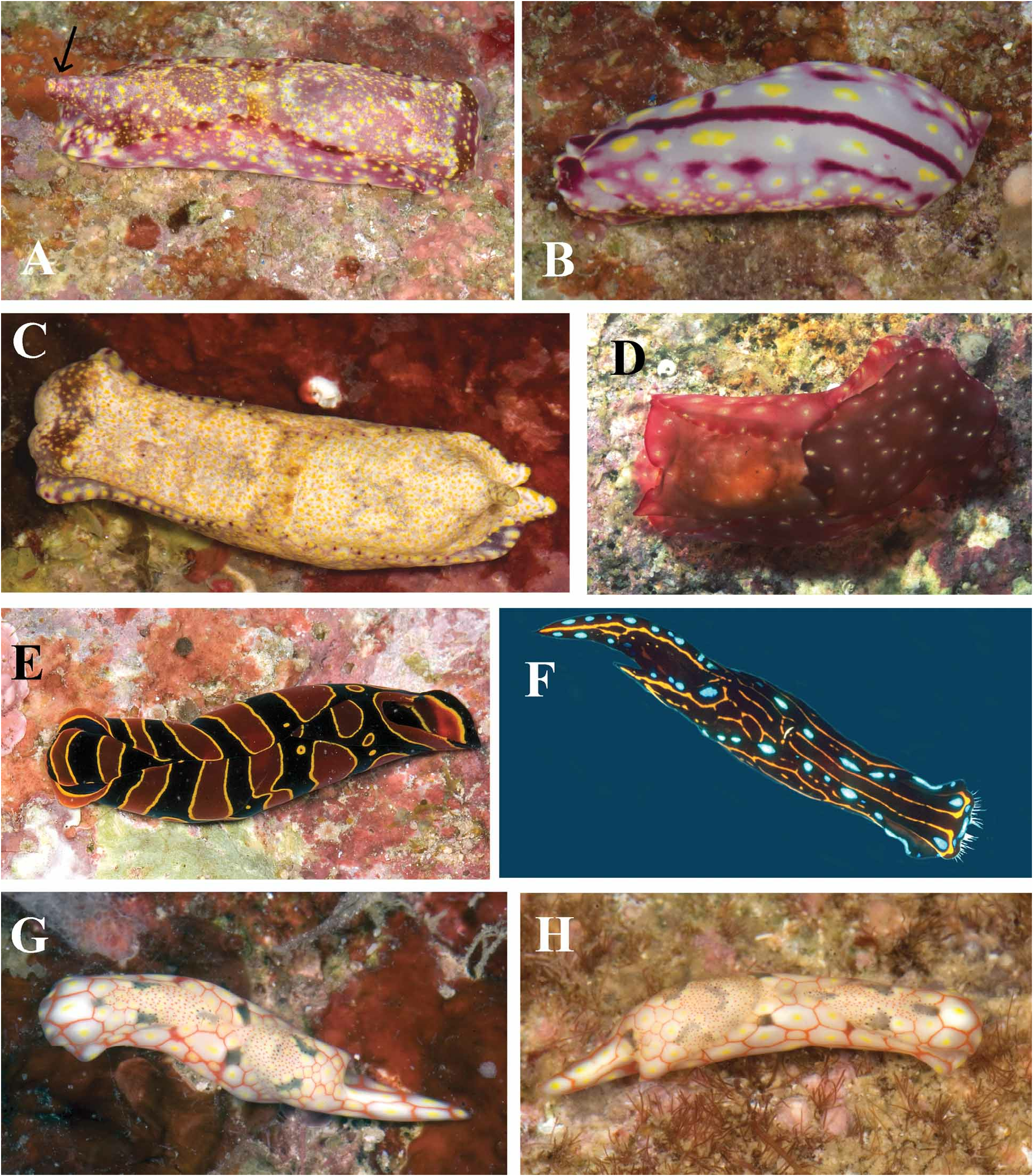
Description. External morphology. The living animals are 9 to 25 mm in length and 5–12 mm wide. The general body color of the living animal ( Fig. 1A View FIGURE 1 ) is light pinkish to wine red with maroon with darker red pigment on the anterior portion of the head, along the parapodial margins and at the base of the posterior projection of the posterior shield. The entire dorsal and lateral surfaces of the body are ornamented with scattered irregular yellow spots and a few whitish patches. Specimens from the Marshall Islands ( Johnson 2008) are much paler with dorsal and lateral whitish pigment and a few maroon spots on the head and parapodial margins. Small yellow to orange spots are also present over the body surface. The ventral surface of Philippine animals ( Fig. 1B View FIGURE 1 ) is pale pink to wine red with a midventral longitudinal maroon line and two additional more lateral lines that may be interrupted as maroon spots. The foot has a few large yellow spots scattered over the ventral surface. A similar pattern is found on the foot of the Marshall Island specimens and also those from Red Sea ( Koretz 2005a).
Living animals are elongate, and wide. The anterior end of the cephalic shield is blunt and quadrangular. The cephalic shield is roughly rectangular and terminates posteriorly with a short, rounded papilla. The posterior shield is slightly rounded anteriorly and terminates in a medial, elongate conical posterior projection that is well-elevated from the base of the shield. The two lateral posterior lobes of the posterior shield are short and simply rounded. The parapodia are relatively short, leaving most of the cephalic and posterior shields visible. The gill is simply plicate consisting of 13 primary folds and is situated ventrally on the right posterior end of the animal.
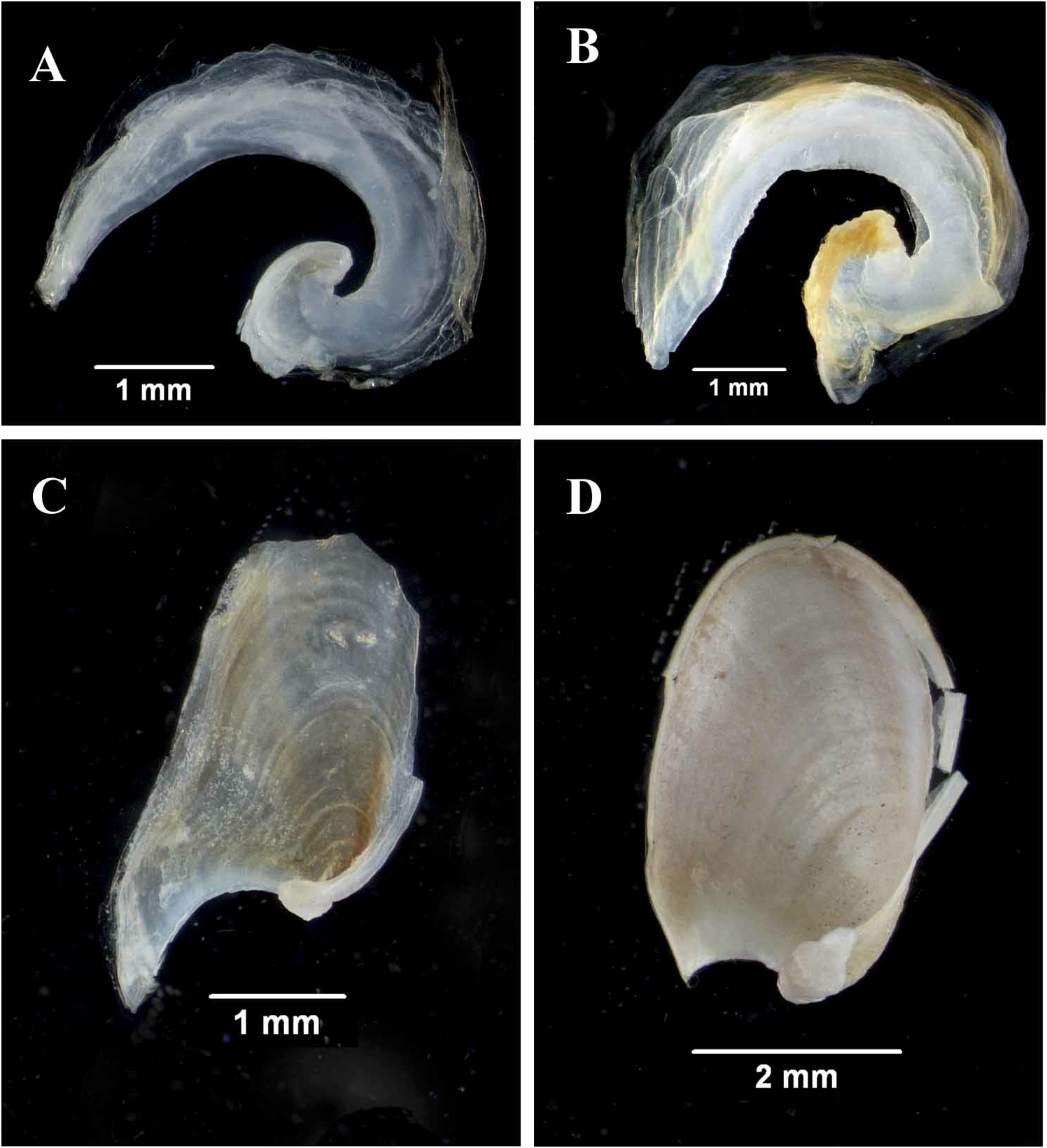
Shell ( Fig. 2A View FIGURE 2 , 3A View FIGURE 3 ). The shell is relatively thickly calcified and consists of a narrow band that occupies the posterior extreme of the animal. There is a membranous periostracum that is slightly more extensive anteriorly than the calcified portion. The area at the base of the shell near the protoconch is more thickly calcified that the rest of the shell.
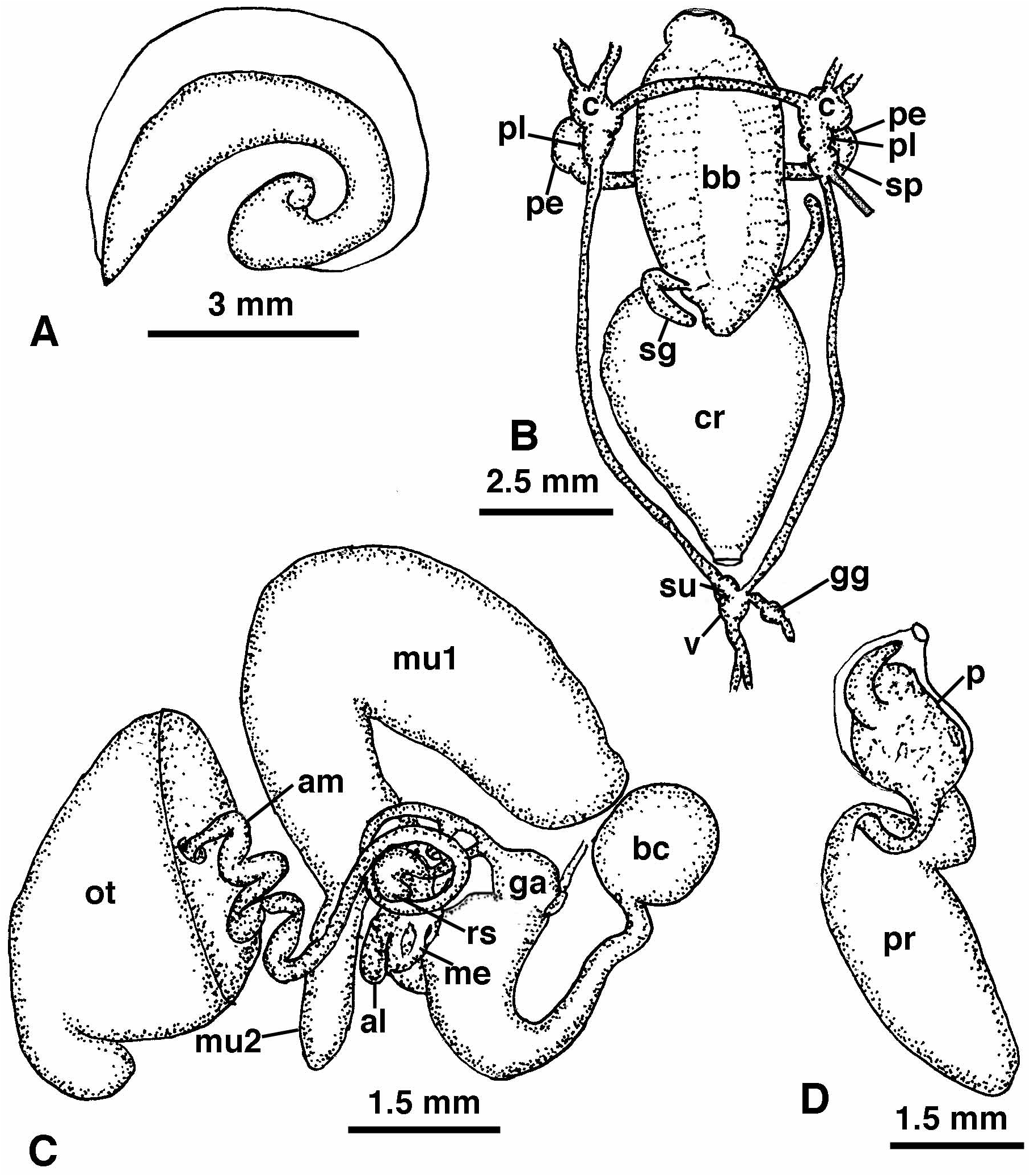
Digestive system ( Fig. 3B View FIGURE 3 ). The buccal mass is large, highly muscularized and slightly elongating posteriorly and occupies the body cavity for the entire length of the cephalic shield. The buccal bulb entirely lacks any vestige of a radula. There is a large ventral oral gland and small dorsal oral glands were indistinct. At the posterior end of the buccal mass, near the junction with the crop, is a pair of elongate salivary glands. The crop is large and saccate, wider than the buccal bulb. The crop narrows posteriorly and enters the digestive gland. The intestine emerges from the right side of the digestive gland and terminates near the posterior end of the body near the base of the gill.
Central Nervous System ( Fig. 3B View FIGURE 3 ): The circumesophageal nerve ring consists of paired cerebral, pedal and pleural ganglia and a single supraintestinal ganglion on the right side. The cerebral and pedal commissures are both elongate with well-separated respective ganglia. Immediately adjacent and posterior to the right pleural ganglion is the supraintestinal ganglion. From its posterior end is the right branch of the visceral loop and the osphradial nerve. The two lateral branches of the visceral loop join posteriorly at the posterior ganglia. The left visceral loop enters the subintestinal ganglion, while the right lateral nerve enters the visceral ganglion. The visceral ganglion is larger than the subintestinal ganglion. From the visceral ganglion is the genital nerve, which has a distinct genital ganglion.
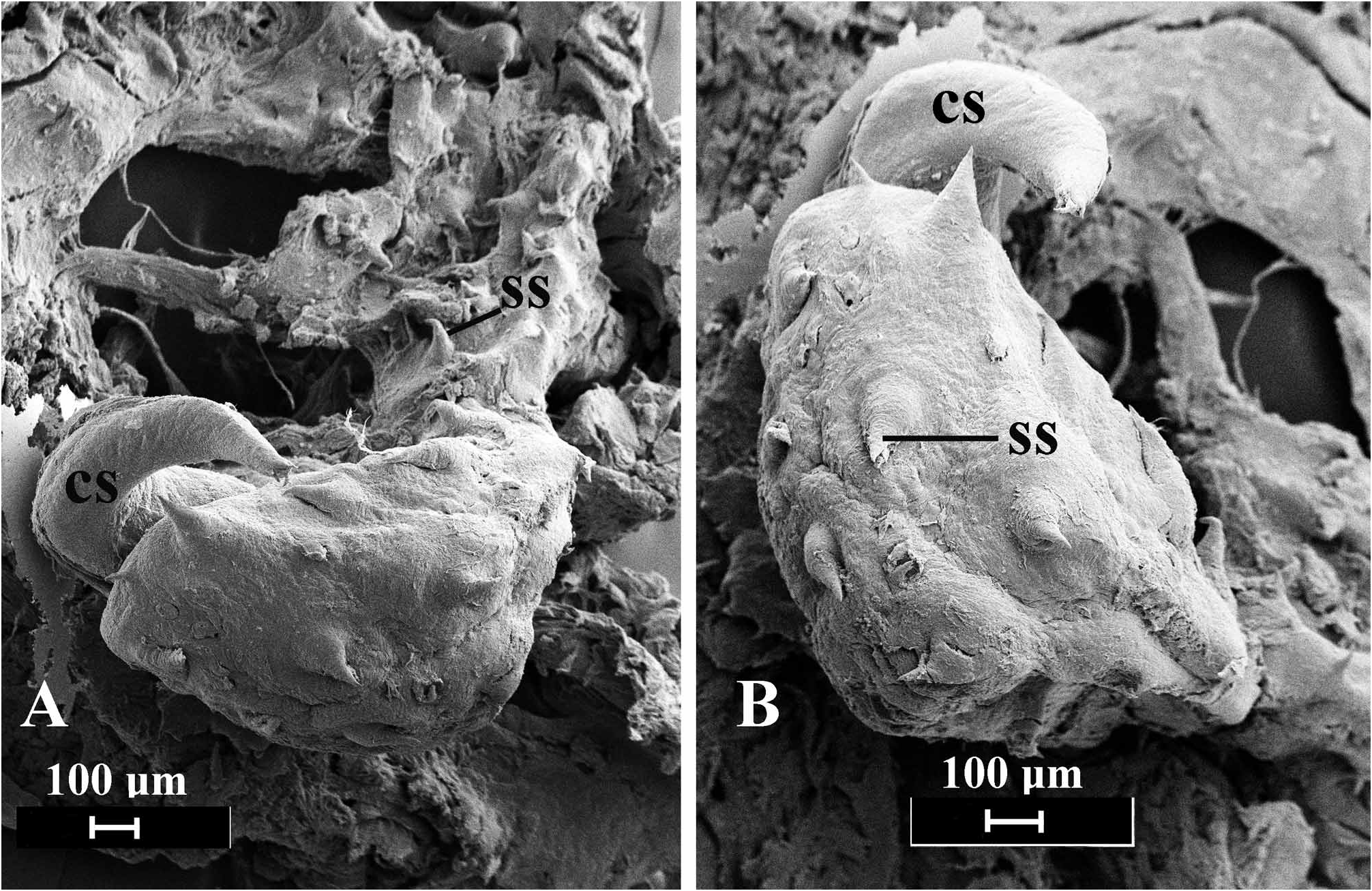
Reproductive System ( Fig. 3C–D View FIGURE 3 ). The arrangement of reproductive organs is essentially monaulic (as discussed by Gosliner 1994) but with a single branch of the hermaphroditic duct to the albumen and membrane glands ( Fig. 3C View FIGURE 3 ). From the large ovotestis, which is intermingled with the digestive gland, emerges the convoluted ampulla. The ampulla narrows into the hermaphroditic duct, which curves around the receptaculum seminis and enters the short, coiled albumen and membrane glands by means of a single duct. The larger mucous gland is bilobed with a massive primary lobe and smaller secondary one. The hermaphroditic duct then joins the duct of the receptaculum seminis and continues to the genital atrium, where it joins the duct of the bursa copulatrix. The bursa is large and spherical. Its duct is narrow where it joins the bursa and widens until its widest portion at the genital atrium. From the genital atrium the open, ciliated sperm groove leads to the cephalic penis. The penis ( Fig. 3D View FIGURE 3 ) consists of a penial sac and a lobate prostate gland that is joined to the penial sac by a narrow duct. Within the penial sac is a large penial papilla ( Fig. 4 View FIGURE 4 ) that is covered by a scattered series of large spines that are distributed over the surface of the penis and along its right base. Extending from the right surface of the penial papilla is a large curved, sickle-shaped chitinous spine.
Remarks. The presence of a quadrangular anterior end of the body, a posterior lobe of the cephalic shield, a large muscular bulb, the single branch of the hermaphroditic duct to the albumen and membrane glands, a bilobed mucous gland are all characteristics of species of Philinopsis .
In his review of Philinopsis, Rudman (1972a) indicated there were two distinct groups of species. Members of the first group are characterized by having a large quadrangular head, a bulbous buccal mass and a specialized penis with a basal elongate penial papilla and a short prostate ( Marcus & Marcus 1967: fig. 12). Members of the second group have a rounded head with an elevated bulbous region (that resembles the front end of a Boeing 747) with prominent eyes visible at the base of the bulb, an elongate, tubular buccal bulb and a simple penis. Gosliner (1980) suggested that Aglaja depicta ( Renier, 1807) , placed by Rudman in Aglaja , had attributes more similar to the members of the quadrangular-headed species of Philinopsis and tranfered it to this genus. While the remainder of the anatomy of P. depicta is similar to the members of the quadrangular-headed group, it has a simple penis. Philinopsis falciphallus has a quadrangular head but differs from other members of this group in having a conical posterior lobe to the posterior shield. It also has a bulbous muscular buccal bulb. The penis of P. falciphallus is unique among described aglajids in several respects. It is the only species that has the prostate separated from the penial bulb by a narrow duct. It is also the only Philinopsis with an armed penis. Not only is the penis armed, but it has a series of spines on the papilla and a large curved chitinous spine. A few other aglajid species have an armed penis. Melanochlamys diomedea (Bergh) ( Rudman 1972a) and M. barryi Gosliner ( Gosliner 1990: fig. 14) have a single apical penial stylet. Melanochlamys papillata Gosliner ( Gosliner 1990: fig. 8) and Aglaja ocelligera (Bergh) ( Rudman 1974: fig. 13) have a penis with numerous papillae on the penial papilla. None of those structures resemble the elaborate armature present in P. falciphallus .
A preliminary molecular phylogenetic analysis (Gatdula et al. personal communication) using the 16S mitochondrial gene fragment has this species clustered with group of Philinopsis species that have a tubular buccal mass rather than those that have a bulbous buccal mass. This is despite the fact that P. falciphallus has a bulbous buccal bulb. More detailed analysis is required using multiple genes, but preliminary data clearly indicate that this species is nested within the clade of Philinopsis species.
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Philinopsis falciphallus
| Gosliner, Terrence 2011 |
Philinopsis sp. 5
| Gosliner, T. M. & Behrens, D. & Valdes, A. 2008: 42 |