
Phallangothelphusa zenuorum, Campos & Alexander, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4851.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:F32CF559-B473-4DDE-B986-3ED23FE8E2E1 |
|
DOI |
https://doi.org/10.5281/zenodo.4408158 |
|
persistent identifier |
https://treatment.plazi.org/id/B32BA7B9-06F6-44F4-9BCC-5740C6E02667 |
|
taxon LSID |
lsid:zoobank.org:act:B32BA7B9-06F6-44F4-9BCC-5740C6E02667 |
|
treatment provided by |
Plazi |
|
scientific name |
Phallangothelphusa zenuorum |
| status |
sp. nov. |
Phallangothelphusa zenuorum View in CoL , new species
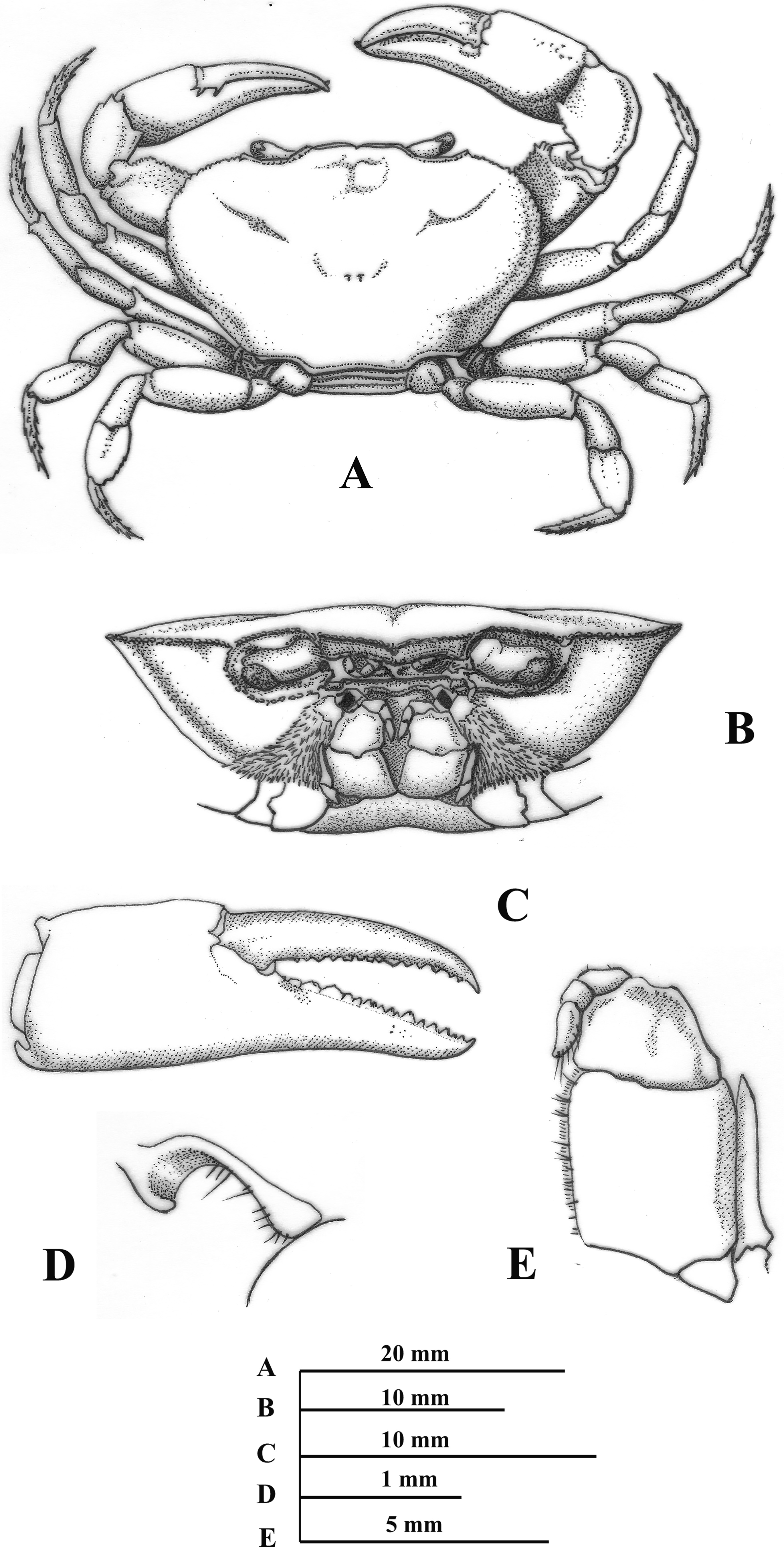
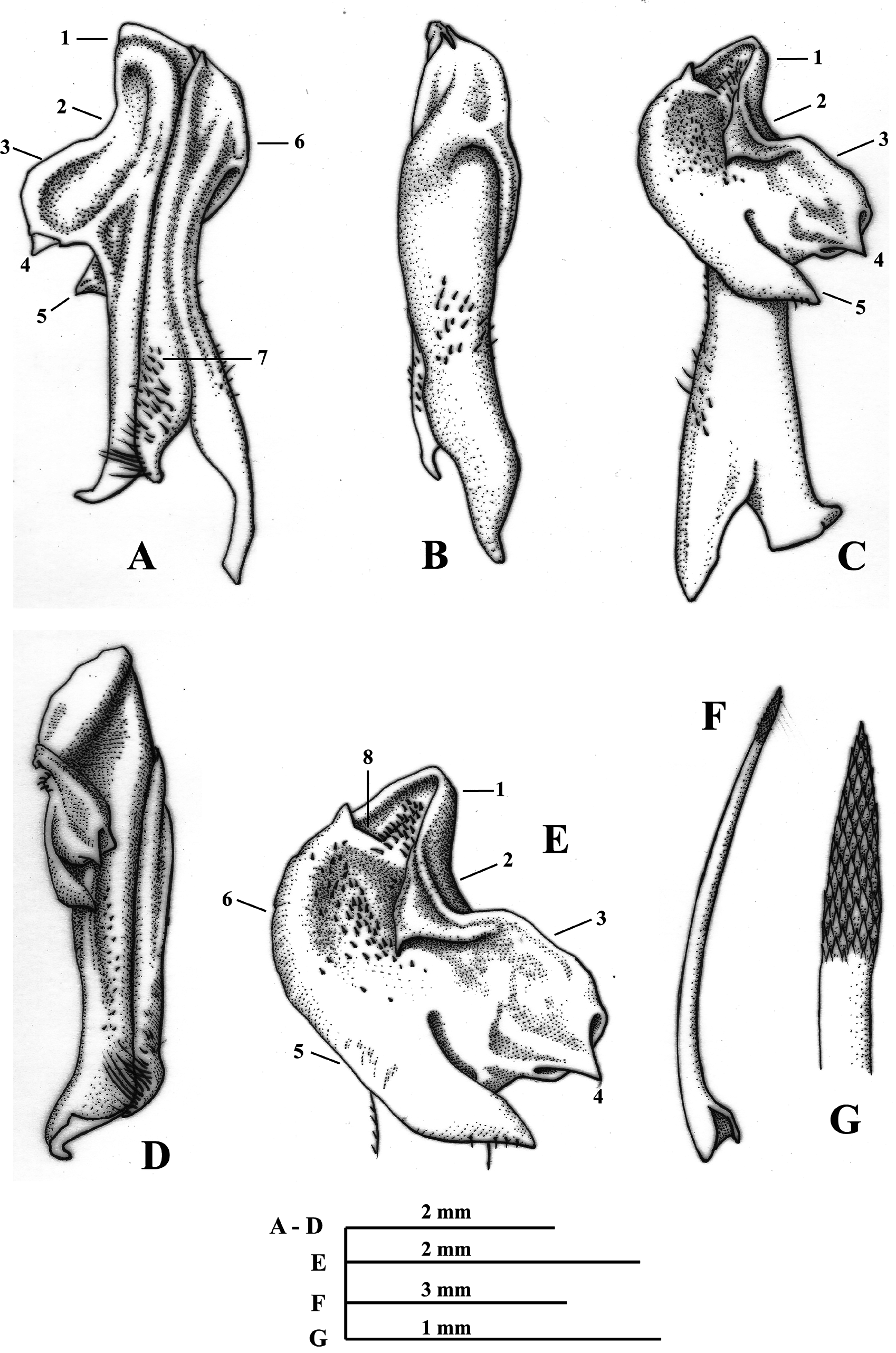
( Figs. 1 View FIGURE 1 , 2 View FIGURE 2 )
Material examined. Holotype: male (cl 16.3 mm, cw 28.0 mm), ICN-MHN-CR 3478. Colombia, Antioquia Department, Municipio El Bagre, Vereda Los Almendros, Los Almendros stream, elevation 196 m, 7°27’38.8”N, 74°40’18.4”W, 25 Oct 2019, leg. M. Alexander. GoogleMaps Paratypes: 1 male (cl 14.9 mm, cw 24.8 mm), 1 female (cl 15.0 mm, cw 35.4 mm), 1 juvenile female (cl 12.5 mm, cw 21.6 mm), ICN-MHN-CR 3479. Same locality data as for holotype GoogleMaps .
Diagnosis. Mesial process of first male gonopod formed by two projections (distal, proximal) former subtriangular, second one oblong with basal transverse slit through which an acute spines is projected, distal and proximal prouections separated by narrow curvature; lateral process protuberant, nearly semicircular with a conspicuous acute spine distally, tip upwards; lateral process as long as proximal projection of mesial process; apex outline nearly subtriangular in distal view, with internal margin convex and external margin concave, bearing numerous, conspicuous spinules; espermatic channel with rows of conspicuous spinules. Second male gonopod with endopod concave, rounded and covered by rows of scale-like spinules distally. Third maxilliped with exopod as long as outer margin of endopodal ischium.
Description of holotype. Carapace ( Fig. 1A View FIGURE 1 ) with sinuous, narrow, deep cervical grooves, becoming indistinct towards margins of carapace; antero-lateral margins with deep notch behind outer orbital angle, with 8 papillae before cervical grooves; lateral margins with approximately 16 papilliform teeth, diminishing in size posteriorly; postfrontal lobes high, oval, delimited anteriorly by transverse depression; median groove deep; surface of carapace between front and postfrontal lobes inclined anteriorly and towards mid-line; front high, slightly excavated, upper border convex with middle depression in dorsal view, marked with row of papillae, lower margin strongly sinuous in frontal view, visible in dorsal view, conspicuously thickened, marked with row of papillae; upper, lower orbital margins each fringed with tubercles ( Fig. 1B View FIGURE 1 ); dorsal surface of carapace smooth, covered by small papillae, regions demarcated; third maxilliped with deep depression on subdistal external margin of merus, exopod as long as outer margin of endopodal ischium ( Fig. 1E View FIGURE 1 ); orifice of efferent branchial channel open, nearly ovate with slight extension of the lateral lobe of the epistome ( Fig. 1D View FIGURE 1 ).
First pereopods heterochelous ( Fig. 1A View FIGURE 1 ); right cheliped larger than left, merus with 3 crests as follows: upper crest with rows of tubercles of different sizes, internal lower crest with 2 tight rows of large tubercles, diminishing in size proximally, external lower crest with row of papillae; carpus with blunt subdistal spine, follows by 4 rounded tubercles, decreasing in size proximally; palms of both chelipeds smooth, swollen, with rows of papillae on external, internal margin, fingers of chelae with rows of dark tubercles, not gaping when closed, tips crossing when closed ( Fig. 1C View FIGURE 1 ). Walking legs (second to fifth pereopods) slender ( Fig. 1A View FIGURE 1 ), dactyli each with 5 longitudinal rows of large spines diminishing in size proximally, arrangement of spines on dactylus of left second pereopod as follows: anterolateral, anteroventral rows each with 6 spines, external row with 7 spines, postero-ventral, posterolateral rows each with 5 spines.
First male gonopod straight, wide; caudal lobe wide basally, decreasing in size to apex, ending acute in caudal view, basally, covered by rows of conspicuous setae; prominent ridge basal to beyond mid portion, running parallel to caudal lobe; lateral side concave basally, middle wide constriction and rounded protuberance from subdistal portion to apex; mesial process formed by two projections (distal, proximal) separated by narrow curvature in between, distal projection subtriangular, rounded distally in caudal view, basal portion folded over distal surface, ending in acute spine, directed downwards, proximal projection oblong, recurved meso-caudally with basal transverse slit through which an acute spines is projected, this spines is originated on cephalic surface of proximal projection ( Figs. 2A View FIGURE 2 , C-E); lateral process protuberant, near semicircular, wide, rounded basally and decreasing in size distally, inwardly turned, directed latero-mesial, with a conspicuous acute spine distally, tip upwards; lateral process as long as proximal projection of mesial process and separated each other by deep gap; apex outline nearly subtriangular in distal view, with internal margin convex and external margin concave, bearing numerous, conspicuous spinules; espermatic channel with rows of conspicuous spinules ( Fig. 2C, E View FIGURE 2 ). Second male gonopod with endopod concave, rounded and covered by rows of scale-like spinules distally ( Fig. 2F, G View FIGURE 2 ).
Color. The freshly-alcohol preserved holotype is brown (near Mars Brown, 223A) with light specks (near Cinnamon, 123A) on the dorsal side of the carapace. The walking legs dorsally brown (near Verona Brown, 223B), ventrally light brown (near Mikado Brown, 121C). The chelae brown dorsally (near Mars Brown, 223A), light brown ventrally (near Buff, 124). The ventral surface of the carapace is light brown (near Sayal Brown, 223C).
Habitat. The specimens were collected by hand under stones and leaf litter of a secondary forest stream. The stream is surrounded by a gallery forest and characterized by clear waters, with stones, sandy-clay soil, limestone substrate and leaf litter at the margins. The freshwater Ecoregion of the locality corresponds to the Lower Cauca, which is characterized by alluvial plains of tectonic depressions and narrow valleys and canyons Mesa et al. (2016). This region has abundant wetlands and associated flood areas that subject to the dynamics of flood pulses, since the hydrographic systems of the Cauca and Nechí rivers converge. The Corrales and El Ocho swamps on the eastern bank of the Nechí River and the foothills of the western flank of the Serranía de San Lucas are of the highest importance due to their extension. These boggy complexes are part of an orographic complex that constitutes one of the Pleistocene refuges known as the Nechí Refuge ( Haffer, 1969), characterized by presenting very high levels of speciation, numerous endemisms and a remarkable richness of species Acosta-Galvis & Restrepo (2001), Constantino & Rodríguez (2009).
In this Ecoregion there are registers of deforestation and legal and illegal mining. This leads to consequences for the quality of the habitat where the species is found. Recently, the Corporación Autónoma Regional del Centro de Antioquia, CORANTIOQUIA, established the Lower Cauca Ecoregion as one of the priority areas to carry out Biodiversity conservation actions, which will contribute to the protection of forests and water sources and consequently the conservation of species.
Etymology. The species in named after the Zenú Indians, in whose territory the species was found.
Remarks. Rodríguez (1982) hypothesized that the shape of the orifice of the efferent branchial channel and the length of the exopod of the third maxilliped are primitive conditions of Phallangothelphusa , and that probably this genus is derived from an ancient stock of Strengeriana Pretzmann, 1971 . For Phallangothelphusa zenuorum n. sp., we find that the exopod is as long as outer margin of endopodal ischium, similar charateristic that is presented in some species of the genus Strengeriana : S. antioquensis Prahl, 1987 , S. cajaensis Campos & Rodríguez, 1993 and S taironae Rodríguez & Campos, 1989 .
In a cladistic study of freshwater crabs of the tribe Strengerianini , belonging to the subfamily Pseudothelphusinae, Rodríguez & Campos (1989) used somatic and gonopodal characters, and selected as out-group Epilobocera Stimpson, 1860 , that is the only genus of the subfamily Epilobocerinae , with distribution in the Antilles. Epilobocera species are characterized by having the exopod of the third maxilliped distinctly overreaching the ischium, the presence of a tooth inside the efferent branchial channel and the absence of a field of spines in the gonopod Rodríguez (1982), Rogers et al. (2020).
For the cladistic study Phallangothelphusa dispar , the only species known of this genus at that time, was excluded of the analysis, because the closure of the efferent branchial channel is achieved by the elongation of the epistome, not homologous to the advancement of the jugal angle of the others genera belonging to Strengerianini . However, Rodríguez & Campos (1989), Fig. 5, based on homologies in the gonopods, postulate that Phallangothelphusa dispar could be derived from Strengeriana restrepoi Rodríguez, 1980 .
The species of Phallangothelphusa are distributed in upper and middle Magdalena Valley region. The discovery of Phallangothelphusa zenuorum n. sp. extends the range of the genus to the foothills of the Serranía de San Lucas, Nechí river basin, that is tributary of the Cauca river, and the elevation ranging from 196 to 1500 m above sea level.
Phallangothelphusa zenuorum n. sp. is most similar to Phallangothelphusa martensis Cardona & Campos 2012 (see Campos 2014, figs. 222A–E). The main distinguishing features between the two species are in the first male gonopod: (1) the lateral side in P. zenuorum n. sp. has a middle wide constriction and rounded protuberance from subdistal portion to apex in caudal view ( Fig. 2A View FIGURE 2 ), whereas in P. martensis it is narrower near subdistal portion, hourglass-like, followed by sinuous border to apex ( Campos 2014: fig. 222B); (2) the distal projection of the mesial process in P. zenuorum n. sp. presents the basal portion folded over distal surface, ending in acute spine in distal view ( Fig. 2C, E View FIGURE 2 ), whereas in P. martensis it is ending in acute spine basally, and it is not folded over distal surface ( Campos 2014: figs. 222D, E); (3) the proximal projection of the mesial process in P. zenuorum n. sp. is oblong with basal transverse slit through which an acute spines is projected in caudal view, this spines is originated on cephalic surface of proximal projection ( Fig. 2A, C, E View FIGURE 2 ), whereas in P. martensis it is rectangular, with a notch instead of a transverse slit on basal portion and devoided of spine ( Campos 2014: figs. 222B, D, E); (4) the two projections of the mesial process (distal, proximal) in P. zenuorum n. sp. are separated by narrow curvature in between ( Fig. 2A, C, E View FIGURE 2 ), whereas in P. martensis this is widest ( Campos 2014: figs. 222B, D, E); (5) in P. zenuorum n. sp., the lateral process is protuberant, wide, rounded basally and decreasing in size distally, inwardly turned, directed latero-mesial, and shows a conspicuous acute spine distally, with tip upwards ( Fig. 2A, C, E View FIGURE 2 ), whereas in P. martensis it is sinuous basally, parallel to axis of gonopod, aproximately 60% of length, the remaining 40% is projected mesially, the basal portion is covered by rows of spinules ( Campos 2014: figs. 222B, D, E); (6) the apex outline in P. zenuorum n. sp. is nearly subtriangular in distal view, with internal margin convex and external margin concave ( Fig. 2C, E View FIGURE 2 ), whereas in P. martensis it is oval in distal view and near flat distally in caudal view ( Campos 2014: figs. 222D, E). The endopod of the second gonopod in P. zenuorum n. sp. is concave, rounded and covered by rows of scale-like spinules distally ( Fig. 2F, G View FIGURE 2 ), whereas in P. martensis it is slightly sinuous with internal portion nearly flat, and covered by rows of spinules distally ( Cardona & Campos, 2012, fig. 2F, G)
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Strengerianini |
|
Genus |