
Phaenocora evelinae Marcus, 1946
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3889.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:67896601-F3C6-44F2-A237-78120C8EA5DB |
|
DOI |
https://doi.org/10.5281/zenodo.5660127 |
|
persistent identifier |
https://treatment.plazi.org/id/CF039A58-FFC6-C532-17C4-0CD7E7EAFA9A |
|
treatment provided by |
Plazi |
|
scientific name |
Phaenocora evelinae Marcus, 1946 |
| status |
|
Phaenocora evelinae Marcus, 1946 View in CoL
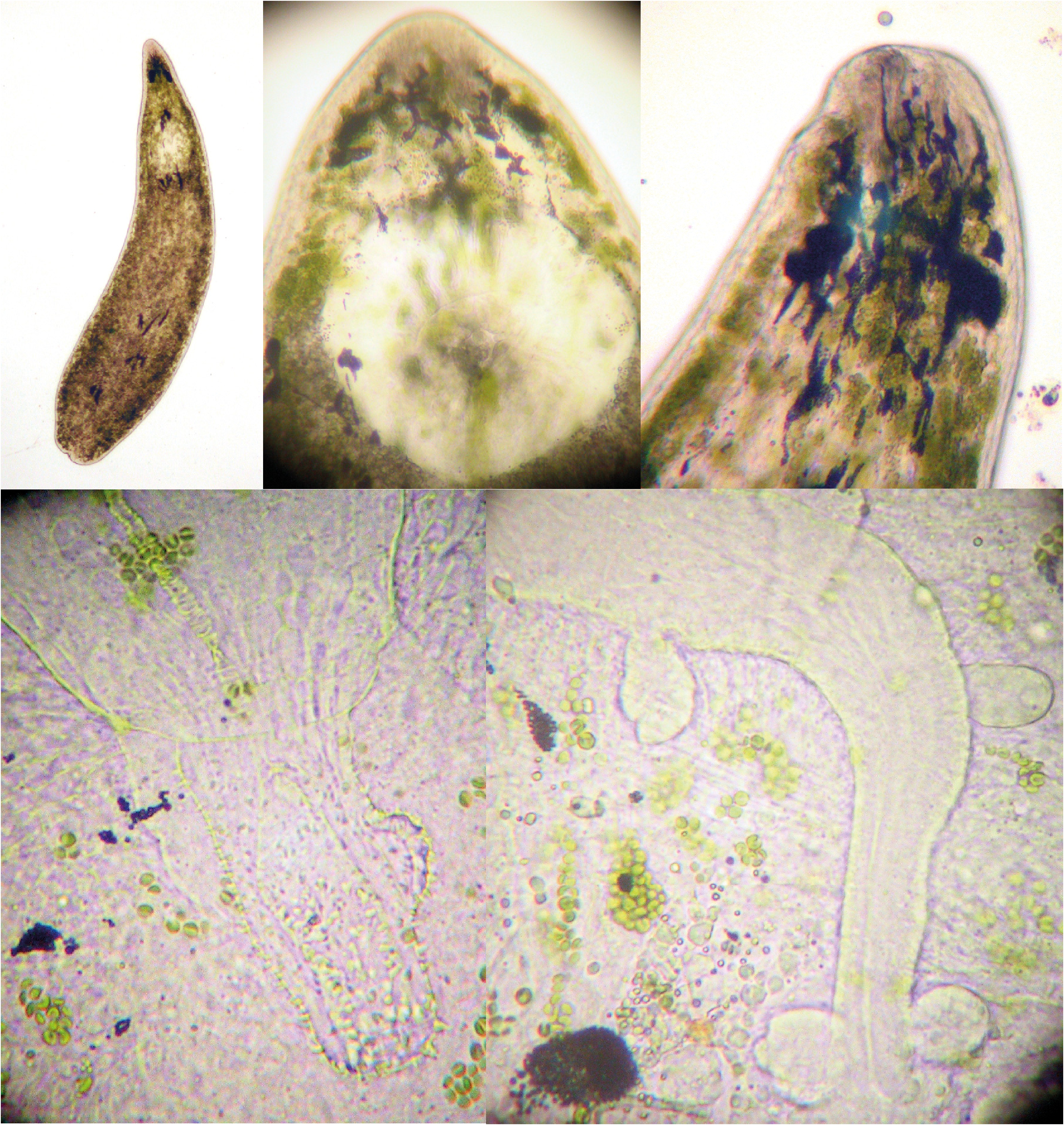
( Figs 1 View FIGURE 1 A, 5A–F, 13C)
Phaenocora evelinae Marcus 1946: 70 View in CoL , 72–78, 80–82, 164–166, Figs 84, 88–91, plate 16 Fig. 2; Marcus & Marcus 1959: 18 –20; Noreña et al. 2004: 234.
Known distribution: near the bridge de Jaguaré between the roots of Eichhornia crassipes View in CoL (Water hyacinth); 1 km south of the Butantan institute (Butantâ, São Paulo, Brazil), in stagnant water, a swamp near the river Pinheiros ( Marcus 1946).
New localities: São Sebastião do Caí, close to Feliz and Montenegro (Rio Grande do Sul, Brasil; 29°36’21,51”S; 51°21’59,66”W), submerged grasses, 19 June 2010.
Material examined: The syntypes: one slide with 15 whole-mounted specimens ( SMNH no. 97006).
Five sagitally-, two horizontally- and one transversely-sectioned specimens from São Sebastião do Caí (sections of eight specimens located on eleven slides: HU nos VI.3.25–VI.3.35). Two whole mounts from São Sebastião do Caí (HU nos VI.3.36–VI.3.37).
Diagnosis: Animals up to 2 mm long. With white eyes and zoochlorellae. Body pigmentation might be present, then situated at level of the anterior part of the pharynx. Male copulatory organ of the duplex-type IIIA. Female genital system of the EVELINAE - type. Distal part of the female genital canal with a diverticulum. Intestinal bursa strongly lobed.
Descriptive notes: The studied animals are about 1 mm long (measured on sagitally-sectioned specimens). The body is rounded anteriorly and ends in a tail ( Fig. 1 View FIGURE 1 A). The eyes have a white appearance under reflective light and black under transmissive light. Zoochlorellae are numerous, which makes the study of the internal organs on live animals difficult.
The inferior genital atrium ( Figs 5 View FIGURE 5 E–F: iga) is lined with a cellular, nucleated epithelium that bears cilia. The superior genital atrium ( Figs 5 View FIGURE 5 E–F: sga) bears the inner lobes of the pear-shaped appendages ( Figs 5 View FIGURE 5 E–F: psl1). Small basophilic glands are attached to the outer lobes of the pear-shaped appendages (not shown on figures).
The male copulatory organ ( Figs 5 View FIGURE 5 B–E) is of the duplex-type IIIA. At least three muscular layers surround this copulatory organ. The innermost muscle layer ( Fig. 5 View FIGURE 5 E: cm i) belongs to the inner muscular septum (see GENERAL MORPHOLOGY section). The other two belong to the outer muscular septum and (probably) consist of an inner circular muscle layer ( Fig. 5 View FIGURE 5 E: cm o) and an outer longitudinal muscle layer ( Fig. 5 View FIGURE 5 E: lm o). The penis papilla is lined with a thin epithelium without spines and is (probably) surrounded by muscles, some of which appear to be circular, whereas others are apparently longitudinal (muscles not shown on figures).
The female genital system ( Fig. 5 View FIGURE 5 F) includes a valvular apparatus ( Fig. 5 View FIGURE 5 F: va), a burso-intestinal duct ( Fig. 5 View FIGURE 5 F: dbi), a lobular intestinal bursa ( Fig. 5 View FIGURE 5 F: bi) and a female genital canal ( Fig. 5 View FIGURE 5 F: fgc), which distally bears a diverticulum ( Fig. 5 View FIGURE 5 F: div). The oviduct ( Fig. 5 View FIGURE 5 F: od) opens at the place where the female genital canal enters the intestinal bursa. Shell glands ( Fig. 5 View FIGURE 5 E–F: sg) are present, a fact not mentioned by Marcus (1946), although this author mentions the presence of eggs with a shell.
Remarks: Marcus (1946) mentions the presence of a lobular unicellular gland entering the oviduct. We could not find any such gland on our specimens. However, exactly at the place where Marcus (1946) mentions such a gland, the vitelloduct enters the oviduct, which was clearly visible in one of the sectioned specimens (sections of one specimen located on two slides: HU nos VI.3.27–VI.3.28). At this spot, the epithelium of the vitelloduct is very high. Possibly, Marcus (1946) misinterpreted this epithelium as the lobular gland.
We also did not observe the tertiary lobes of the pear-shaped appendages mentioned by Marcus (1946) in the original description. However, the illustrations of Marcus (1946) show that he based the distinction between primary, secondary and tertiary lobes upon the shape of the cells: cells within the primary lobes are curved towards the lumen of the superior genital atrium, whereas cells of the secondary lobes are curved outwards. Gilbert (1935), however, already stated that the shape of cells within the lobes can vary, and based the distinction between primary and secondary lobes upon their relative position compared to the muscle layers surrounding the pear-shaped appendages; primary lobes are inside these muscle layers, whereas secondary lobes are outside. Tertiary lobes, as in P. sulfophila , are clearly separate structures that attach to the secondary lobes as defined by Gilbert (1935). Marcus' (1946) drawings clearly show that his primary and secondary lobes are both within these muscle layers, thus representing the primary lobes of Gilbert (1935), whereas his tertiary lobes are outside of it and hence are secondary lobes in Gilbert's (1935) terminology. True tertiary lobes as observed in P. sulfophila are therefore absent in P. evelinae .
Whether the diverticulum of the female genital canal is always single and always at the left side (as stated by Marcus 1946) is not certain. In some of our sectioned specimens (most obvious in HU no. VI.3.26) we have the strong impression that there actually are two diverticula, occurring on opposite sides of the female genital canal. Although the original description contains many details that can only be obtained by the study of sectioned specimens, the only material remaining from that description consists of 15 whole-mounted specimens, all on one slide, representing the syntypes. As these whole mounted specimens do not show any of the diagnostic characters to identify the species, we refrain from designating a lectotype. It is our intention to request the Commission to set these syntypes aside under its plenary power for the designation of a sagitally-sectioned specimen (HU no. VI.3.25) as neotype (ICZN 1999: according to Art. 75.5).

| SMNH |
Saskatchewan Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
SubPhylum |
Rhabditophora |
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Phaenocorinae |
|
Genus |
Phaenocora evelinae Marcus, 1946
| Houben, Albrecht M., Steenkiste, Niels Van & Artois, Tom J. 2014 |
Phaenocora evelinae
| Norena 2004: 234 |
| Marcus 1959: 18 |
| Marcus 1946: 70 |