
Peltophryne armata, Landestoy & Turner & Marion & Hedges, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4403.3.6 |
|
publication LSID |
lsid:zoobank.org:pub:AEABA77E-FABE-4A50-B211-80E6BB8FCE8B |
|
DOI |
https://doi.org/10.5281/zenodo.5951974 |
|
persistent identifier |
https://treatment.plazi.org/id/CF6487DD-FFAE-FFA9-B9B6-FC5E6AF80FC8 |
|
treatment provided by |
Plazi |
|
scientific name |
Peltophryne armata |
| status |
sp. nov. |
Peltophryne armata View in CoL sp. nov.
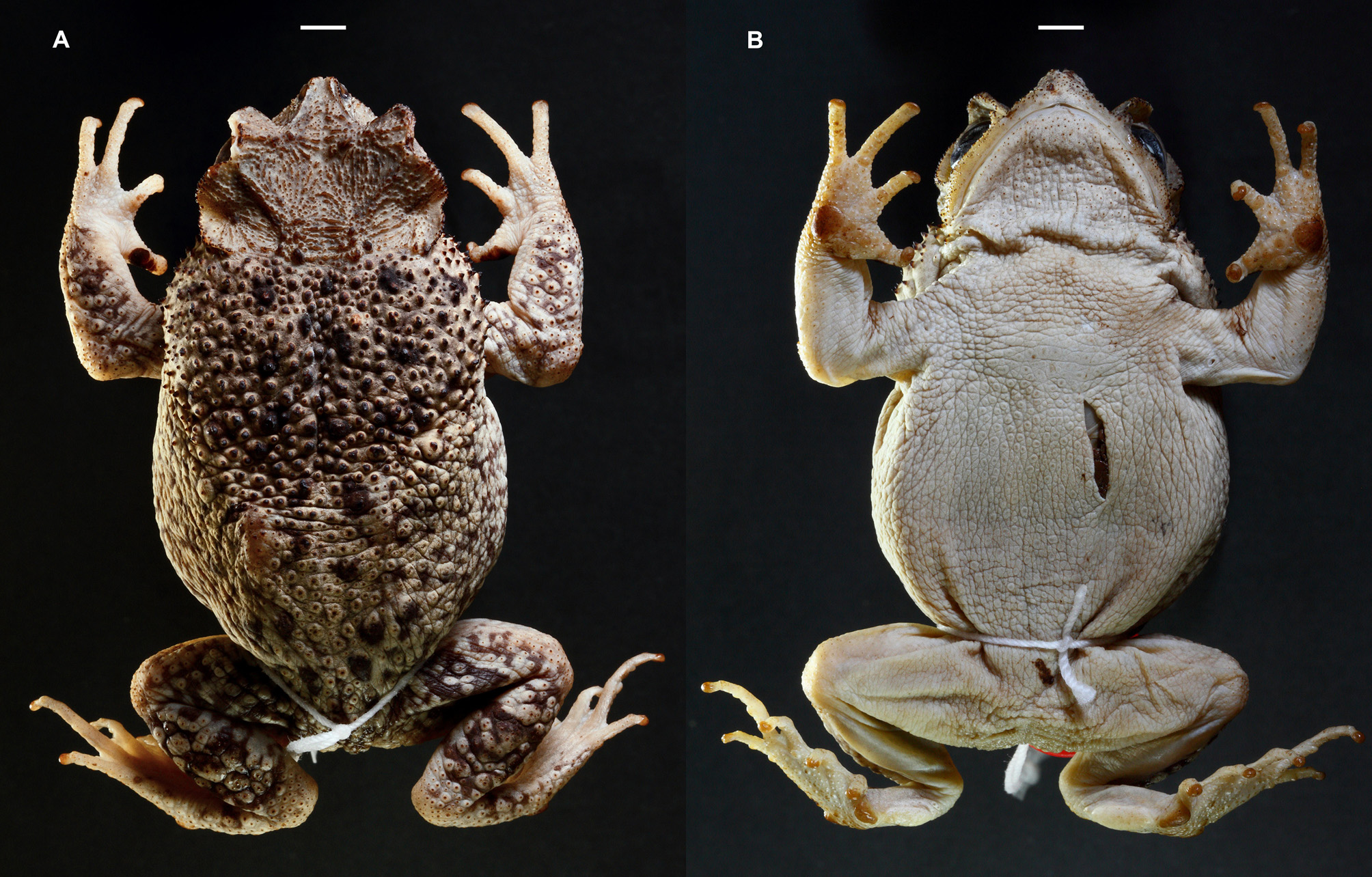
Holotype: MCZ A-149839 ( Fig. 3 View FIGURE3 ), an adult male with SVL of 76.8 mm (weight in life 35.2g), from 1.6 km SSW of Las Mercedes , Sierra de Bahoruco, 345 m elevation, Pedernales Province, Dominican Republic, collected by M. Landestoy, R. Ortíz and Y. Corona on the night of 23 August 2014 .
Paratypes: (n=9; 7 males, 2 females) MCZ A-149840 (female), MCZ A-149841, MNHNSD 23.1375 View Materials , collected by M. Landestoy and R. Ortíz on the 22 August 2014, 1 km SSW Las Mercedes, on the southern slopes of the Sierra de Bahoruco, Pedernales Province ; MCZ A-149842, MNHNSD23.1376–78 , same data as the holotype, collected by M. Landestoy, R. Ortíz and Y . Corona; MCZ A-149843, MNHNSD23.1379 View Materials (female), same locality as former, on 31 August 2014, by M. Landestoy and R. Ortíz.
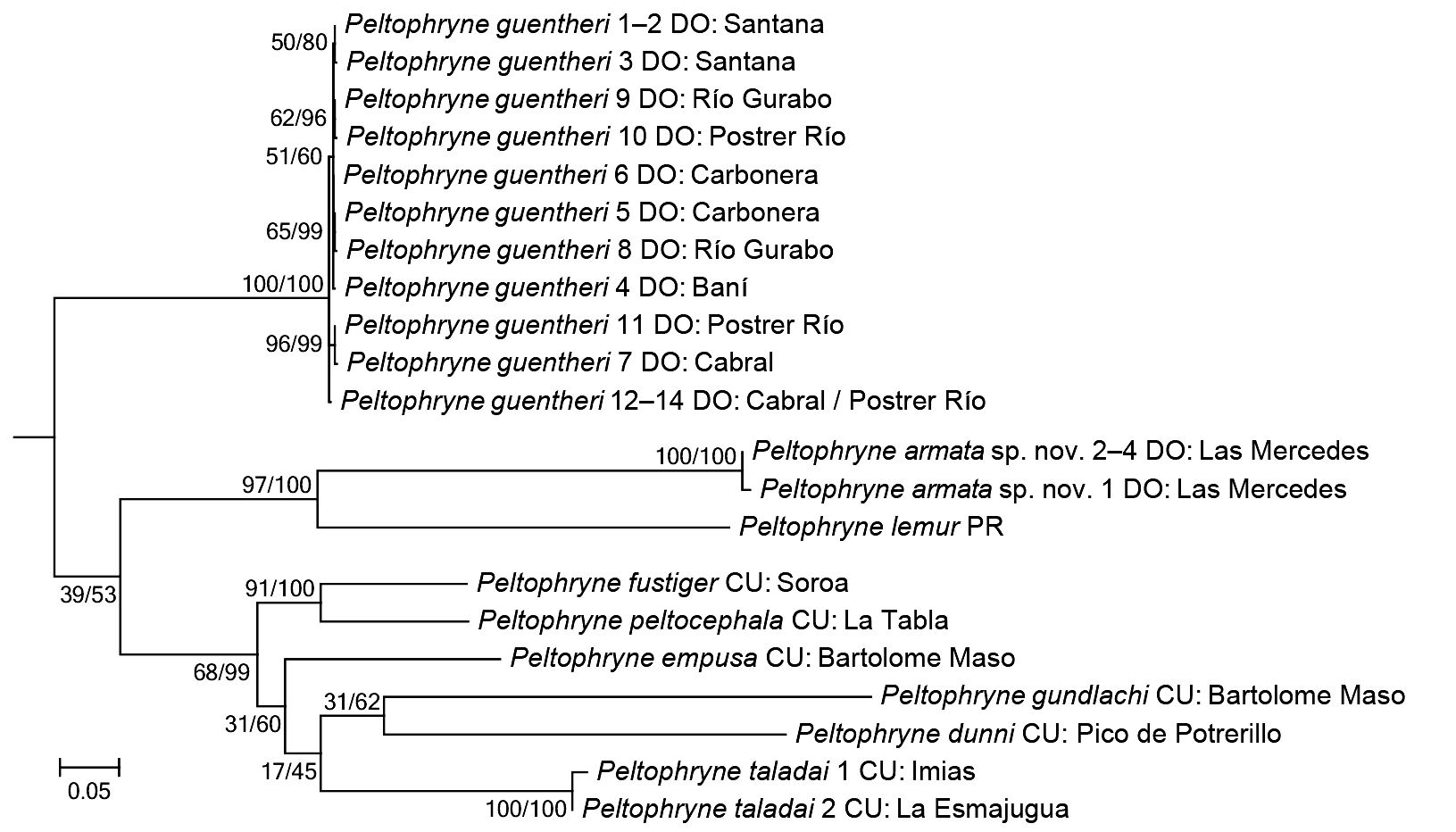
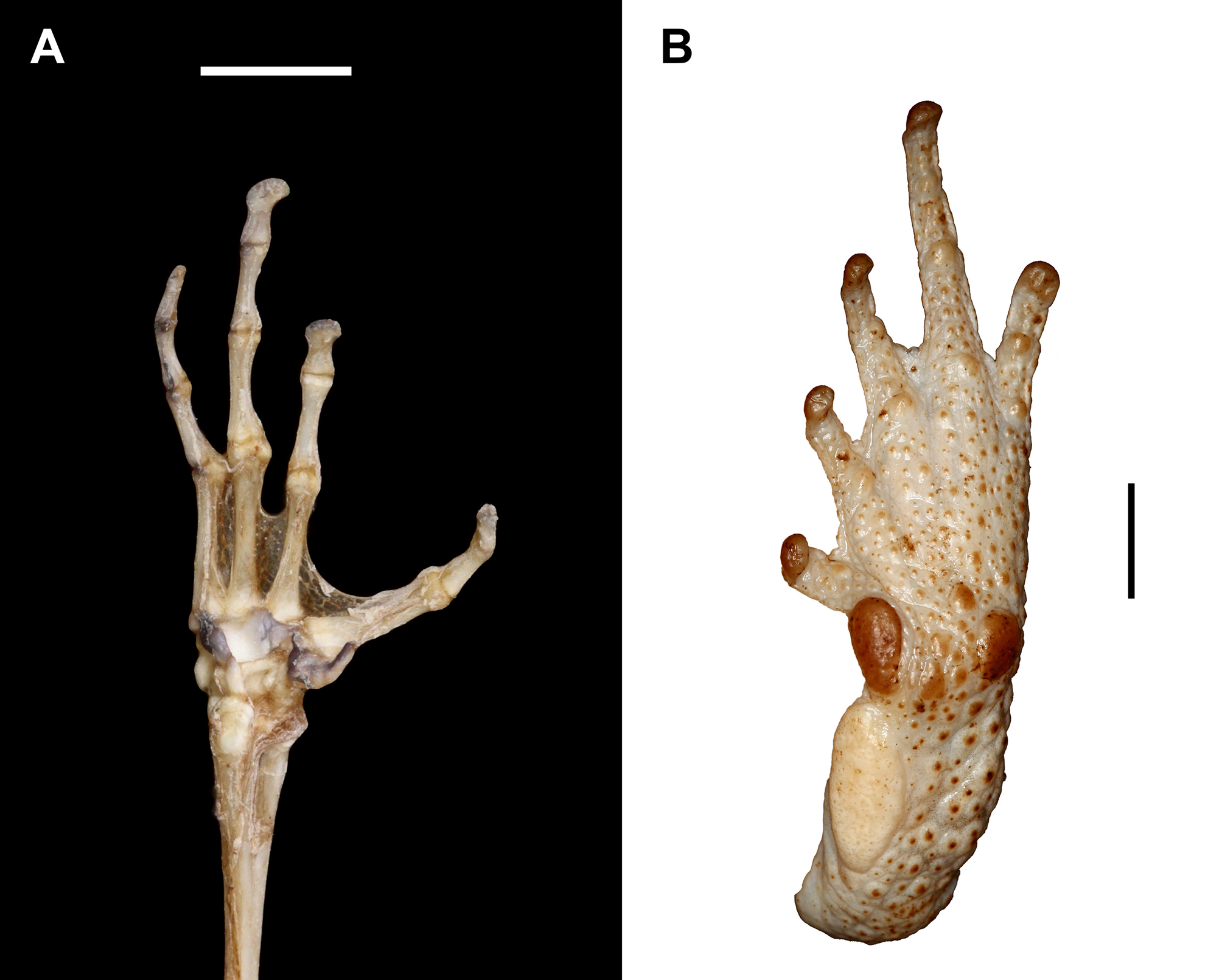
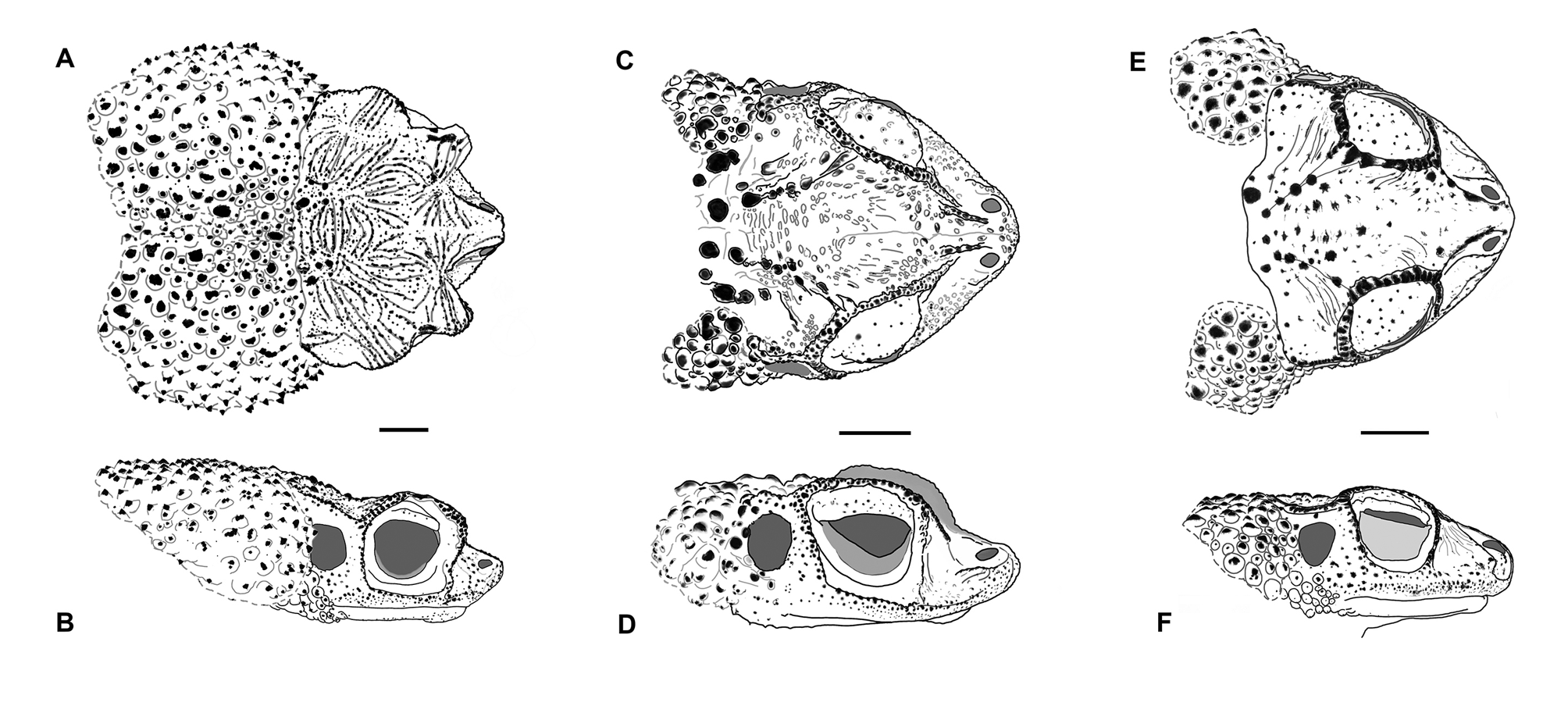
Diagnosis. A medium-sized species of the genus Peltophryne (SVL in males to 78 mm, in females to 85.7 mm), possessing characteristics of the genus: T-shaped terminal phalanges and discrete glandular tubercles on tarsus ( Fig. 4A–B View FIGURE 4 , respectively; sensu Pramuk 2002), and placed within the genus in a molecular phylogeny ( Fig. 2 View FIGURE 2 ). It is distinguished from all other congeners by three unique characters: hypertrophic cephalic crests (supraorbital, postorbital, supratympanic, pretympanic and preorbital crests, the first two crests extended outwards largely covering eyelids and orbits in dorsal view; Fig. 5A–B View FIGURE5 ), hypertrophic parotoid glands (longitudinally, transversely and obliquely massive) converging middorsally, and snout with a notched tip (distal ends of canthal crests create a gap in between). It is most closely related in a molecular phylogeny to the Puerto Rican species P. lemur , with which it shares a depressed head (in profile), very low or absent infraorbital crest (merged with the maxillary crest), a protrusive, slightly upturned snout, large orbits, and a long, complex (Type II) advertisement call. Beyond its unique characters, the new species also differs from that species by the geometry of crests on the dorsal surface of the head (intersection of postorbital and supraorbital crests forming obtuse angles—acute with canthal—versus these crests being nearly continuous and straight to slightly concave in P. lemur ; Fig. 5C View FIGURE5 ), by having the head rounded (versus subtriangular in P. lemur ), by having the parotoid glands with spinose keratinized tubercles (versus subrounded keratinized tubercles in P. lemur ), by having feet basally webbed (versus strongly webbed in P. lemur ), and in having a large vocal sac (versus small to moderate in P. lemur ).
Peltophryne armata is less closely related to Hispaniolan species and it is not known to be sympatric with any of its congeners from that island. Nonetheless, besides its unique characters it can also be distinguished from its geographic neighbor on Hispaniola, P. guentheri , by the obtuse angles at intersection of supra- and postorbital crests (versus right angles in P. guentheri ; Fig. 5E–F View FIGURE5 ), rounded head shape in dorsal view (versus subtriangular in P. guentheri ), a Type II call (versus short Type I call with evenly spaced, rapid rising simple pulses in P. guentheri ), the presence of a thick and pad-like tarsal fold (versus a narrow and sharp flap-like ridge in P. guentheri ), in the basal, short foot webbing (versus large foot webs in P. guentheri ), and a whitish or cream-colored throat in adults of both sexes (versus dark olive pigmentation in males of P. guentheri ).
Description of the holotype. MCZ A-149839 ( Fig. 3 View FIGURE3 ), measurements in Table 1. Habitus stout and head broader than long (HL1 87.4% of HW1), nearly rounded in dorsal view, relatively depressed (dorsoventrally) in profile. Supraorbital and postorbital crests very long, directed anterolaterally, gradually elevated from interorbital and parietal areas (creating a concavity in those areas), extended far outwards (in dorsal view) covering most of the eyelids, the intersection of both crests creating a rather obtuse angle, and a high, broad arch well above the eye (in profile), its edges with granular keratinization, the surface of each of those crests bearing several (4–6) transverse keratinized ridges arranged parallel to each other. Supratympanic crest long, extended laterally and elevated from its junction with postorbital crest, covering tympana and curved inward at posterior end (dorsal view) with its outer edges bent upward, the crest oriented diagonally in profile with its lower end joining pretympanic crest. Pretympanic crest prominent, directed anterolaterally. Infraorbital crest undistinguished or absent, overlapped by a prominent maxillary crest. Preorbital crest long, with a median notch, continuous to supraorbital crest but transversed by 4–6 prominent keratinized ridges, and ventrally in contact with and protruding transversely to maxillary crest, its lower end sinuate, bent anteriorly before joining the maxillary crest. Canthal crest prominent, sharp and slightly concave, the distal ends of canthals leaving a gap or notch in between. The convergence of the canthal, supraorbital and preorbital crests forming acute angles, the intersection of the latter two crests projecting anteriorly, sharply convexed in a horn-like form (in dorsal view). Subnasal crest low. Parietal crest relatively low and an indentation between its posterior end and the supratympanic crest. All cephalic crests beaded with moderate level of granular keratinization. Upper eyelid with a prominent row of glandular keratin-tipped tubercles on outer margin. Orbits large, directed anterolaterally. Snout rather short, protruding, semi-truncate in dorsal view (with a notch at tip), sub-acuminate in profile. Nostrils near the tip of snout, directed dorsolaterally. Interorbital and parietal areas heavily ornamented with numerous keratinized ridges and rugosities. Posterior of head with high bony conical tubercles. Loreal area slightly concave, with spinose keratinized tubercles scattered around snout and below tympanum down to the angle of jaw. Tympanum distinct, nearly rounded, its posterior partially overhung by parotoid gland. Parotoid glands very large and spinose (multiple keratin tips), elongate longitudinally and very broad transversely, almost touching each other middorsally (with a narrow groove in between them which widens anteriorly towards the head), and obliquely large, extending down below the level of the angle of jaw where distinct, but indistinct posteromedially, breaking into large keratinized tubercles and warts dispersed middorsally, decreasing in size, prominence and level of keratinization dorsolaterally and towards the cloaca. Flanks with low and small tubercles. Forelimbs with moderate keratin-tipped rounded tubercles, decreasing in size and prominence towards elbows and hands. Hindlimbs rather short. Dorsal surface of shanks with enlarged rounded glandular tubercles each with multiple keratin tips, smaller tubercles on thighs. Tarsus with moderate rounded tubercles on outer margin, its inner margin with an elongate and thick tarsal fold, pad-like ( Fig. 4B View FIGURE 4 ). Skin of venter granular, the granules low, keratin-tipped anteromedially. Throat with higher keratin-tipped granules. Margins of jaw with a row of low keratin tips.
All palmar tubercles keratinized. Thenar tubercle small, rounded, nearly one-third the size of the oval palmar tubercle. Median subarticular tubercles in fingers divided or bifid, proximals single and rounded to subconical. Supernumerary tubercles rounded and low. Fingers with well-developed lateral fringe, unwebbed, their relative lengths III>IV>I=II.
All plantar tubercles keratinized. Inner metatarsal tubercle large, ovoid. Outer metatarsal tubercle oval, twothirds the size of the inner metatarsal tubercle. Subarticular tubercles of toes single, rounded and low. Supernumerary tubercles low. Tarsal fold elliptical and thick, pad-like ( Fig. 4B View FIGURE 4 ). Toes with well-developed lateral fringe, basally webbed (distinct between toes II-III and III-IV), their relative lengths IV>III>V>II>I.
Color in life. ( Fig. 6B View FIGURE 6 ) Dorsal ground color (including dorsal surfaces of limbs and eyelids) bright yellow with several black spots on dorsum, arranged paravertebrally with fewer, smaller spots scattered in parotoid glands. The head is darker yellow, with shades of dark brown in the interorbital and parietal areas, and some dark spots in its posterior among the bony conical tubercles. There is a subtle greenish cast behind head, at the middorsal intersection of parotoid glands, and on thighs. Diffuse and disrupted charcoal-gray vermiculations in dorsolateral and posterior areas of body, and on forelimbs, with the vermiculations darker, continuous and broader on hindlimbs, with three transverse bands across thighs and shanks. The large and glandular tubercles and warts on hindlimbs are predominantly yellow, encircled by very dark pigmentation, similar to those tubercles on forelimbs.
Flanks have fainter dorsal ground color and pigmentation of the vermiculated pattern, fading into cream-white lateroventrally. Hands and feet are cream-pinkish, with supernumerary tubercles buffy in hand, cream in feet. The venter is unpatterned, cream-white anteriorly, flesh-colored posteriorly (including limbs), with somewhat translucent skin revealing a blue-purplish hue most noticeable lateroventrally, and under shanks and tarsi. All granules and tubercles on venter are cream-white. The chin, the mid-throat and its edges are cream-white finely stippled with light gray, and the vocal sac is cream-white. The iris is greenish-gold, with many black venations in its lower section, less so in its upper section. Pupils are bordered with black extended across the corners of eye.
Color in preservative. Dorsum and limbs light tan, with diffuse charcoal-gray vermiculations, and dark blotches as described above (in life); head of a darker brownish color; venter grayish.
Variation. Cephalic crests slightly variable in extension, especially the supraorbitals, and to a lesser extent, the postorbitals and preorbitals. A female (MCZ A-149840) has significantly shorter crests than those in the rest of the series. Apparent sexual dimorphism: males are smaller (SVL up to 78 mm, versus 85.7 mm in females) and with more developed cephalic crests. Dichromatism: males with bright yellow dorsal ground coloration, females with a more tan yellow to light ochre color. In most males, the intersection of supraorbitals and preorbitals is convex, directed anteriorly in the form of horns in dorsal view; parietal crests generally prominent, inconspicuous in one side in some specimens (MCZ A-149839 and MCZ A-149843), and inconspicuous in both sides in others (MNHNSD-23.1376–77). An elevated, prominent knob is at the posterior edge of the supratympanic crest in MNHNSD-23.1378 and MCZ A-149839. The interorbital area has either continuous or discontinuous ridges and bony tubercles. In some specimens the snout is nearly rounded in profile. The tympanum is oval to nearly rounded in all specimens except in female MCZ A- 149840 in which it is nearly rectangular. Description of coloration and pattern in life of the holotype consistent with rest of males in series. A male (MCZ A-149843) with broader black vermiculations and blotches. Some specimens have single orange spots on the head (MCZ A-149840, MNHNSD- 23.1375). Males turned dark brown (dorsal surfaces) when in concealed places, and changed to a bright yellow when exposed. Venter and throat are pattern-less cream-white in all specimens.
Juveniles. Metamorphs of Peltophryne armata measure ~ 9.5 mm in SVL ( Fig. 7A View FIGURE 7 ), and are distinct from those of Peltophryne guentheri in having a dark chocolate brown dorsal ground color with light salmon-colored blotches in the supra- and post-tympanic areas and in the sacral area (larger blotches) that extend laterally. An interorbital bar of same color extends at times as an inverted T towards the snout. Limbs are predominantly bonecolored with dark brown elbows, knees and dorsal surface of hands with a continuous dark stripe that crosses the thigh, shank, tarsus, and foot. Venter dark to rich brown, freckled with grayish-bone color. Toadlets ( Fig. 7C View FIGURE 7 ) begin acquiring a green coloration replacing the dark brown dorsal ground color from a size of> 20 mm SVL. Venter turns predominantly bone color with light brown to pinkish vermiculations. Hands and feet ivory-yellow, the palmar and plantar bases being brownish, with tubercles bright yellow. The parotoid glands, pointy snout (in profile), and cephalic crests become evident at such size range.
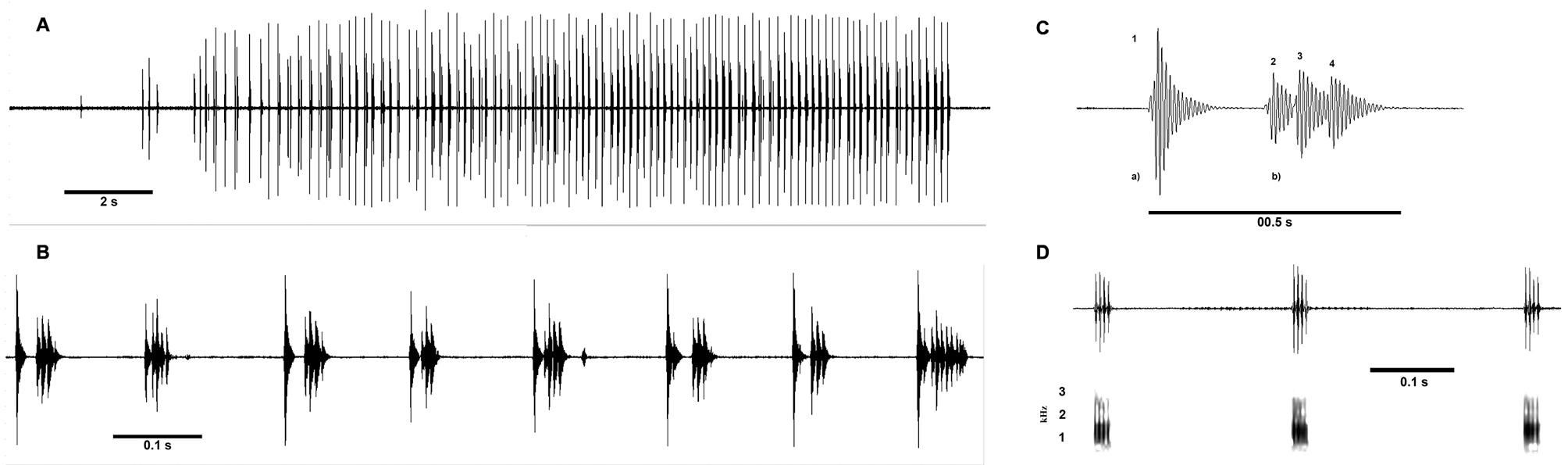
Vocalization. The advertisement call of Peltophryne armata is a series of clucking pulses (range 24–103 pulses, Tab. 2, Fig. 1A–C View FIGURE 1 ). The call sequences (n=12) are composed of trains of pulses, most of which contain subpulses within ( Fig. 1C View FIGURE 1 ), characterized by a pattern of amplitude modulation of the Type II. Call duration range is 5.8– 19.6s (x̄= 16), and can bear 24–103 (x̄= 78.3) pulses per call. Pulses (n=959) can bear 1–10 subpulses (x̄= 4.1). The DF of calls was 1.2 kHz, although single component pulses in the beginning of call have DF of 0.86 kHz, and 5 components of 1.37 kHz (n=1). Intervals between calls range from 41 to 180s (x̄= 93). Bandwidth 1.03–1.55 kHz (x̄= 0.52). 90% bandwidth was measured at ±.52 kHz in the power spectrum of a call -10 dB from the peak. Data are from a recording made of male MCZ A-149843 just before being collected at 2030 h, sitting next to (78 cm) and facing a water hole (already with tadpoles); air temperature was 24.2°C, and relative humidity 80.7%.
The release call ( Fig. 1D View FIGURE 1 ) is a soft, three to four note bark-like staccato (n= 2): “peck peck peck”, of a duration of 0.5– 0.6s; intervals between the pulsatile notes (n= 5) 0.20– 0.28s (x̄= 0.23); subpulses in a note 2–4; dominant frequency 1.31 kHz. When handled or pressed in refugia, individuals are capable of emitting a high-pitched "scream" as their distress call.
Acoustic Characters P. armata sp. nov. n =12 P. guentheri n =12 Distribution ( Fig. 6A View FIGURE 6 ). Known only from the vicinity of the type locality, south of Las Mercedes in the southern slopes of the Sierra de Bahoruco north of the Barahona peninsula, Pedernales Province, bordering both the Sierra de Bahoruco and Jaragua national parks in the karst foothills (330–400 m) of the Sierra de Bahoruco.
Natural history. The habitat of Peltophryne armata sp. nov. is semideciduous forest in the limestone-based southern slopes of the Sierra de Bahoruco. Males call ( Fig. 6B View FIGURE 6 ) next to rain water-filled holes in the limestone of mean dimensions (n=4, with eggs) of 17.2x 32.5 cm in diameter and 21.9 cm in depth, where paired strings of eggs are laid and the larvae develop. One clutch contained 420 eggs. Neither amplexus nor oviposition were observed. Several males vocalized in proximity to cavities in the limestone floor where they retreated when disturbed. Two of these males experienced quick metachrosis, having a bright yellow dorsal ground color when exposed but turning brown once inside such cavities. Males MCZ A-149839 and MCZ A-149843 called in concealment from holes. Males MNHNSD-23.1376 and MCZ A-149843 performed phragmosis while hiding in such cavities, and when pushed, used their heads as shields and inflated their bodies. One individual (MNHNSD-23.1376) emitted a distress call when removal was attempted; those males and others also observed entered their respective cavities by walking backwards. All males (except MCZ A-149841 and MNHNSD-23.1375) had supraorbital crests that were injured (bleeding), missing some of the keratin and dermal tissue. This was apparently caused by the sharp surfaces of the karst substrate. The highly developed crests likely serve as a protection from the harsh environment as well as from potential predators. Individuals inflate their bodies when handled and are capable of a sudden release of air as a distress call. Skin secretions, besides those expelled from the distinctive glandular areas, are spread over dorsal surface (including eyelids) and are of a yellowish-ochre color.
On the second night of the first documented breeding event, 23 August 2014, there were approximately 18 calling males, five of which were collected. Other anurans observed and frequently heard at the type locality were the direct-developing frogs (terraranans) Eleutherodactylus alcoae and E. pictissimus . No other larvae-rearing anuran was observed using the breeding microhabitat at the same time as Peltophryne armata , although during an earlier visit (February 2013) to the area, tadpoles of the Hispaniolan Laughing Treefrog ( Osteopilus dominicensis ) were observed in a large hole nearby to the north of the type locality within a disturbed area, and that species has been heard calling at Las Mercedes village.
Toadlets larger than 20 mm ( Fig. 7C View FIGURE 7 ) become greenish in dorsal ground color, retaining the large light blotches. This pattern matches the surrounding substrate, composed mostly of a limestone-based floor with scattered debris and the abundant herbaceous plant Callisia repens ( Commelinaceae ; Fig. 7B View FIGURE 7 ). Similarly, a species of pyrgomorphid grasshopper found near the type locality resembles the plant leaves in shape and color, probably for protection from predators ( Perez-Gelabert 2000; Perez-Gelabert pers. comm.).
Tadpole. Lots MNHNSD 23.1425-A–T, 23.1426-A–D, 23.1427-A–D; Fig. 8A–E View FIGURE 8 . Works on tadpoles of Hispaniola's native bufonids were lacking just until recently, when Incháustegui et al. (2014) offered the first descriptions of the tadpoles of Peltophryne guentheri . Rivero et al. (1980) provided a textual description of the tadpole of the Puerto Rican Crested Toad ( P. lemur ); Díaz & Cádiz (2008) presented tadpole descriptions for Cuban toad species with illustrations of body and mouth morphology, accompanied by a brief account on their biology. Earlier descriptions of Cuban toad tadpoles were for P. longinasus Stejneger ( Ruíz García 1980) , and P. taladai ( Díaz et al. 2000) .
Twenty eight tadpoles of Peltophryne armata in Gosner stages 31–42 were examined. Nineteen measurements (in mm), taken as in Altig & McDiarmid (2006), were made on specimen MNHNSD 23.1427-B, stage 36: 27.5 total length, 9.5 body length, 18.2 tail length, 2.9 tail muscle height at base, 2.6 tail muscle width at base, 2.1 maximum dorsal fin height and 1.7 maximum ventral fin height located 11.2 from body terminus, 6.7 body width located 6 from snout, 5.5 body height located 6.9 from snout, 1.1 eye diameter, 0.4 pupil diameter, 3.2 interorbital distance, 0.4 narial diameter, 1.5 internarial distance, 1.7 snout to naris, 3.2 snout to eye, 6.8 snout to spiracle, 1.4 naris to eye, and 3.2 transverse diameter of oral disc.
Body ovoid (dorsal view), slightly depressed (profile), and somewhat flat ventrally unless eggs have been ingested (globular, see below); eyes dorsal, directed dorsolaterally; snout long, protruding in a shark-like manner (profile); spiracle sinistral; vent tube medial; nostril nearer the eye than snout, of semi-circle ("D") shape, bearing a surrounding rim with a middorsal protuberance; guts thick and not covering most of the belly, easily visible through the largely translucent venter; belly globular after ingestion of eggs; tail musculature dorsally and ventrally convexed; conspicuous pustulose tubercles on dorsum from stage 41.
Oral disc anteroventral, emarginated, with submarginal papillae; a wide dorsal gap, and a very narrow ventral gap; labial tooth row formula 2(2)/3[1] (in most specimens the labial ridge of P-1 is interrupted medially, which may bear hillocks in between containing several teeth); second upper row of teeth (A-2) with a moderately wide median gap; third lower row of teeth (P-3) very short, almost of same length of that of the ventral gap; jaw sheaths keratinized, its edges unserrated; upper jaw with a slight median indentation.
In life, the tadpole of P. armata ( Fig. 8D View FIGURE 8 ) is light brown dorsally and nearly translucent lateroventrally, having a scattering of brown color laterally; dorsum mainly brown with some melanophores and golden iridophores becoming evident at advanced stages (>34), extending from nostrils to dorsal base of tail; venter translucent, with scarce scattered pigmentation; hindlimbs and dorsum with white to salmon color blotches (stages>38); iris very dark brown with some lighter brown venations.
The tadpole can be diagnosed by the following combination of characters: total length (TL) large (30.5 mm); light brown coloration; body slightly depressed; oral disc anteroventral; submarginal papillae present; a gap in P-1 present in most specimens (23 of different stages out of 28); unserrated jaw sheaths; long and pointed shark-like snout (in profile); fins and venter largely translucent; at advanced stages, presence of prominent dorsal tubercles, and dorsum and hindlimbs with large whitish patches. The gap in P-1 and the elongate snout appear to be unique among known tadpoles of Peltophryne .
On Hispaniola, the tadpole of P. armata differs from that of P. guentheri in its larger size (30.5 mm TL), while tadpoles of P. guentheri can reach up to 20.6 mm and have a reticulated dorsal pattern ( Incháustegui et al. 2014). It also differs in having an oral disc in anteroventral position (ventral in P. guentheri ; MALT pers. obs.) and a very short P-3, as well as a short ventral gap (wide P-3 and gap in P. guentheri ; Incháustegui et al. 2014). The tadpole of the introduced Rhinella marina Linnaeus has a mostly black body and tail ( Duellman 1978; Incháustegui et al. 2014). In the tadpole of the Puerto Rican P. lemur the gut is not visible through the belly ( Rivero et al. 1980), being translucent in P. armata . In many aspects, the larval morphology of P. armata is more similar to that of P. florentinoi (no specimens examined in this study, but see Table 3), which is also the most distinctive of all other tadpoles of Cuban Peltophryne ( Díaz & Cádiz 2008) . Reproduction of P. florentinoi takes place in small holes in the karst filled with rainwater ( Díaz & Cádiz 2008), which is the same and only situation in which P. armata’s tadpoles have been observed. The tadpole of P. armata is a facultative oophagous. Larvae (from Gosner stage 26) were observed feeding on conspecific eggs, larger larvae engulfing eggs whole while the smaller rasped upon them; larvae also fed on algae (in situ), and scavenged on conspecifics (ex situ).
Eggs. Eggs with pigmented (melanic) animal pole; measured 1.8 mm of diameter ( P. guentheri eggs measure ~ 1 mm). Egg strings paired, collected in the field on 23 August 2014, from several clutches in slightly different stages. A portion of these eggs were left for hatching, of which tadpoles developed to complete metamorphosis. Suggested common name. Hispaniolan Armoured Toad
Etymology. The specific name is a Latin singular feminine nominative adjective meaning “armored,” in allusion to the highly developed and extended cephalic crests that cover most of the head, and to the massive parotoid glands bearing spinose keratinized tubercles.



| MCZ |
Museum of Comparative Zoology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.