
Parelaphinis umtamvuna, Perissinotto, 2022
|
publication ID |
https://doi.org/ 10.37520/aemnp.2022.004 |
|
publication LSID |
lsid:zoobank.org:pub:0F0B8FD8-76B1-44E0-BEDF-77628E7079EA |
|
DOI |
https://doi.org/10.5281/zenodo.7161043 |
|
persistent identifier |
https://treatment.plazi.org/id/73219E23-FFA4-FF9C-14BD-F99AFE9FF82C |
|
treatment provided by |
Felipe |
|
scientific name |
Parelaphinis umtamvuna |
| status |
sp. nov. |
Parelaphinis umtamvuna sp. nov.
( Figs 44–52 View Figs 44–52 , 62 View Figs 62–65. 62–63 )
Type material. Hඈඅඈඍඒඉൾ: J, South Africa, KZN, Umtamvuna NR, 24 Oct 2004, R Perissinotto & L Clennell ( SANC). Pൺඋൺඍඒඉൾඌ: 3 JJ 1 ♀, same data ( BMPC); 3 JJ 2 ♀♀, ibidem, 30 Oct 2004, R Perissinotto & L Clennell ( BMCS, RPGS); 1 J 1 ♀, ibidem, 8 Dec 2004, R Perissinotto & L Clennell ( TGMF); 1J 3♀♀, ibidem, 1 Oct 2005, R Perissinotto & L Clennell ( BMCS, RPGS); 3JJ 3♀♀, ibidem, 6 Oct 2011, R Perissinotto & L Clennell ( RPGS, TGMF); 1 J 1 ♀, ibidem, 17 Oct 2012, R Perissinotto & L Clennell ( RPGS); 1 J, ibidem, 20 Nov 2012, R Perissinotto & L Clennell ( TMSA); 1 ♀, ibidem, 10 Dec 2012, R Perissinotto & L Clennell ( RPGS); 1♀, ibidem, 10 Nov 2013, R Perissinotto & L Clennell ( RPGS); 1♀, ibidem, 27 Nov 2013, R Perissinotto & L Clennell ( RPGS); 1 ♀, ibidem, 6 Dec 2013, R Perissinotto & L Clennell ( TMSA).
Description of holotype male ( Figs 44–51 View Figs 44–52 ). Body. Black, velutinous, with dark grey maculation spread across entire surface, turning pink to brick-red in some areas of pronotum and elytra as result of preservation; with moderately dense round sculpture on head, becoming more scattered and horse-shoe shaped on pronotal declivities but very sparse and horse-shoe to groove-shaped on pronotal disc, scutellum and elytra; dorsum virtually glabrous, with very sparse and short light setae on head surface, pronotal and elytral declivities but lacking completely on pronotal and elytral disc as well as scutellum ( Figs 44, 46, 47 View Figs 44–52 ).
Size. Total length = 11.3 mm; maximum width = 6.4 mm.
Head. Black with extensive grey tomentum across entire surface, except around apical sinuation; clypeus deeply concave with extremely narrow indentation at apex and lateral margins smoothly rounded; all margins sharply upturned and reflexed; entire surface covered in regularly spaced round punctures, but sculpture becoming more dense and irregular around clypeal apex and on margins; few sparse and extremely short pale-yellow setae distributed along entire surface, becoming longer and denser on ocular canthus; antenna black to dark brown, with club approximately as long as flagellum; thin light setae scattered across flagellum, becoming thicker and denser on pedicel ( Fig. 47 View Figs 44–52 ).
Pronotum. Black and velutinous, with grey (turning pink to brick red upon preservation) tomentose areas vaguely forming three coarse dotted longitudinal lines across disc and continuous lining on lateral margins, becoming most expanded just above postero-lateral corners; without any tubercle or ridge in apical area; with rather scattered and shallow round to horseshoe punctures on declivities, virtually disappearing on disc; few, extremely short light setae visible only along lateral margins and declivities; all margins smoothly rounded with minor angulation only on antero-lateral deflections ( Figs 44, 46 View Figs 44–52 ).
Scutellum. Black, velutinous and glabrous, without any trace of tomentose maculation; large but shallow horseshoe punctures equally scattered along entire perimeter, but absent in central part of disc; broadly equilateral triangular in shape, with acute apex and lateral margins inwardly arcuate; lateral grooves moderately wide and deep along entire margins ( Fig. 44 View Figs 44–52 ).
Elytron. Black and velutinous, with extensive grey maculation spread across entire surface (turning pink to brick-red in some areas due to preservation); first, third and fifth elytral costae moderately well developed and clearly visible; humeral callus protruding outwards and subhumeral arch deeply sinuate; posterior margin remarkably linear and without projections or upturning at apex; narrow and shallow horseshoe to grooved sculpture regularly distributed along basal half of striae and on antero-lateral declivities, becoming smaller and sparser on postero-lateral and apical declivities; surface generally glabrous, with few scattered minute setae on lateral and apical declivities ( Figs 44, 46 View Figs 44–52 ).
Pygidium. Hemispherical in shape, with smoothly rounded, domed apex and marked depressions on basolateral corners; with pair of larger, dark grey maculae in medio-basal position and two pairs of smaller maculae, first in latero-basal position and second on sides of apical dome; with dense but shallow rugulose sculpture across entire surface, but hidden in areas covered by grey maculae; glabrous on disc, with few fine light-yellow setae scattered along lateral margins and apex ( Fig. 48 View Figs 44–52 ).
Legs. Black and densely sculptured, with brown claws and tips along spinal margins; tarsomeres moderately elongate, with apical ones about 1.5 times longer than preceding units and all claws gently arcuate; tibiae with coarse and irregular sculpture, covered in sparse and short light-yellow setae, becoming longer and denser on inner margins and from pro- to metatibia; protibia tridentate, with two apical teeth equally well developed, but proximal tooth drastically reduced in size; mesotibia with prominent bidentate medio-distal spine and tridentate apical spine, spurs brown and moderately elongate, thin and sharp; metatibia with outer carina bearing one large but blunt medio-distal tooth, spurs brown substantially more elongate than in mesotibia and sharp ( Figs 44–46 View Figs 44–52 ).
Ventral surface. Black and shiny with small grey tomentum spots visible only near lateral margins of abdominal ventrites; covered in dense and long light-yellow to tawny setae from pro- to metasternum, but setae becoming very sparse on metacoxa, central femoral surfaces, lateral margins of abdominal ventrites, and virtually disappearing in mesometasternal region and on disc of abdominal ventrites; mesosternal lobe flattish, smoothly rounded at apex and not protruding either forward or downwards; metasternal surface exhibiting sparse round to horse-shoe punctures with setae emerging at their centre, and wide concavity along entire median sulcus in shape of oblong arrow head; abdominal ventrites with slight concavity at centre ( Fig. 45 View Figs 44–52 ).
Aedeagus. Parameres elongate, with dorsal lobes laterally constrained at centre, expanding then slightly in apical half and smoothly rounded at apex ( Fig. 49 View Figs 44–52 ); apical area bending remarkably downwards, with dense and fine long pubescence extending posteriorly to about one third of total length of ventral lobes ( Figs 50, 51 View Figs 44–52 ); club-shaped dorsal flat surface drastically reduced and extending towards basal attachment by barely one third of total lobe distance; ventral lobes protruding laterally but visible in dorsal view only in basal third of their total length ( Fig. 49 View Figs 44–52 ).
Variability. This species is generally small, attaining the smallest size among the three species of Parelaphinis , in the range of 9.8–12.2 mm in total length and 5.3–6.9 mm in maximum width. Its body is consistently black and velutinous, with variable degrees of dark grey maculation, which may turn pink to brick-red in some preserved specimens (e.g. Fig. 44 View Figs 44–52 ). In extreme cases, males may lack entirely any maculation, while females always retain some residual spots. Females are also slightly broader than their male counterparts, have wider protibiae but shorter protarsi and antennal clubs ( Fig. 52 View Figs 44–52 ). Their pygidium protrudes beyond the elytral apices and the abdominal ventrites exhibit a moderate convexity. Furthermore, the metatibial spurs of the female are spatulate and the apical edge of its metatibia is only mildly serrated, while in the male these are bluntly acuminate and markedly serrated, respectively.
Differential diagnosis. As indicated above, under the description of P. drakensbergica sp. nov., this species lies at the other extreme of the gradient in morphological characters observed across the genus, exhibiting the narrowest body shape and the most regressive tomentose ornamentation among the three species ( Figs 35 View Figs 35–42 , 44 View Figs 44–52 , 53 View Figs 53–61 ). Unlike in the other two species, the maculation of P. umtamvuna sp. nov. is actually grey, not white, and it can get so dark to the point of becoming undetectable in some extreme male specimens. Other diagnostic characters can be observed at the level of clypeal shape, protibial denticles development and aedeagal parameres morphology. In P. umtamvuna sp. nov., the clypeus shows the most reduced apical sinuation, along with the most rounded lateral margins among the three species of the genus ( Figs 38 View Figs 35–42 , 47 View Figs 44–52 , 56 View Figs 53–61 ). The development of the three protibial denticles in P. umtamvuna sp. nov. is intermediate between the most expanded type exhibited by P. drakensbergica sp. nov. and the least pronounced type seen in P. moesta ( Figs 35 View Figs 35–42 , 44 View Figs 44–52 , 53 View Figs 53–61 ). Finally, the aedeagal parameres of P. umtamvuna sp. nov. possess the longest, and narrowest lobes among the three species. The flat to gently inward sloping club-shaped area of the dorsal lobes in this species extends merely one third behind the apex and is, therefore, the shortest among the three species ( Figs 40 View Figs 35–42 , 49 View Figs 44–52 , 58 View Figs 53–61 ). In frontal view, the apical region of the lobes shows an extreme degree of downward bending, while this is minimal in P. drakensbergica sp. nov. and intermediate in P. moesta ( Figs 42 View Figs 35–42 , 51 View Figs 44–52 , 60 View Figs 53–61 ).
Bionomy. Adult activity has been recorded throughout the Austral spring, with most records in October. Few specimens have been observed in low flight, just above the ground, in the hottest part of the day and only immediately after a substantial rainfall event. Most specimens were retrieved from underground, on rocky terraces where large accumulations of leaf litter with high moisture content had been retained by plant roots or networks of mosses and lichens ( Fig. 62 View Figs 62–65. 62–63 ). Larvae and cocoons collected in the wild were also reared successfully in the laboratory, under environmental controlled conditions and using their natural food substrate, while spraying water at regular weekly intervals until pupation.
Etymology. The species is named after the Umtamvuna River, on the escarpment of which all specimens currently known originate from ( Fig. 64 View Figs 62–65. 62–63 ). The specific epithet is a noun in the nominative singular.
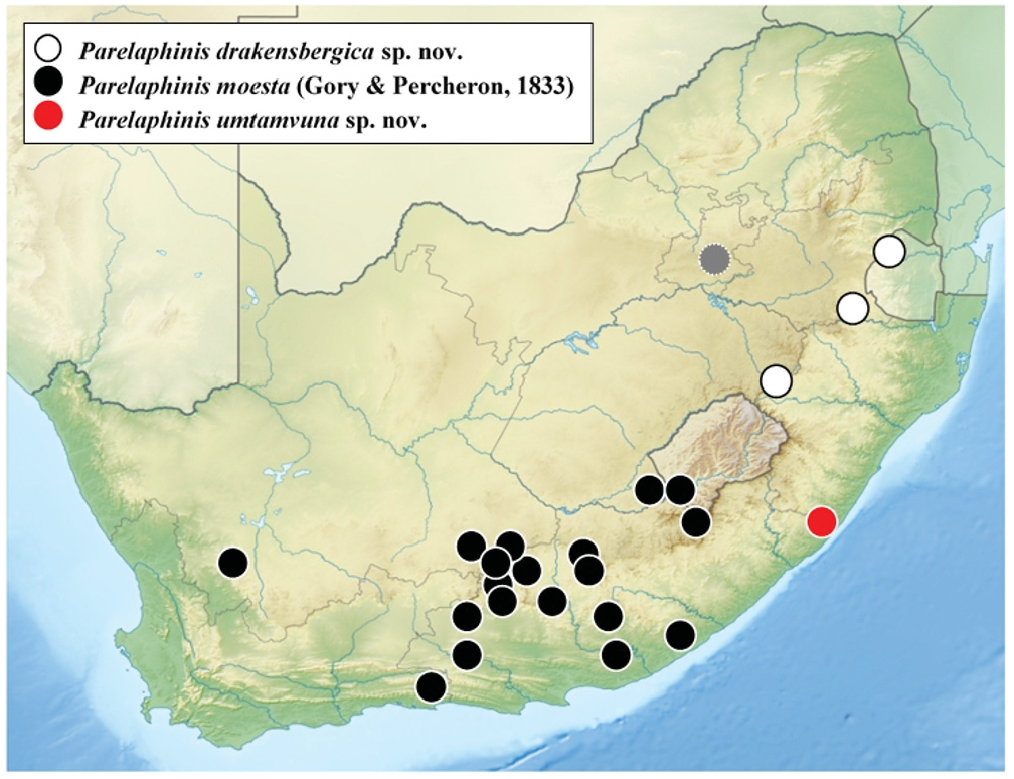
Distribution. This species is so far only known from the upper valley of the Umtamvuna River, on the KwaZulu-Natal side ( Fig. 66 View Fig ). Like the Drakensberg range, this area also experiences a much higher annual rainfall than the western interior of South Africa. Searches in adjacent rivers and tributaries have so far failed to reveal other populations in this province. It is, however, likely that the species occurs also on the Eastern Cape side of this and perhaps other rivers in its vicinities.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.