
Paraonides myriamae Katzmann & Laubier, 1975
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4320.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:878A01B5-C37C-4E46-8631-16927888D8D4 |
|
DOI |
https://doi.org/10.5281/zenodo.6050893 |
|
persistent identifier |
https://treatment.plazi.org/id/5B0CE503-9477-FB56-8482-4FADFCBC7F97 |
|
treatment provided by |
Plazi |
|
scientific name |
Paraonides myriamae Katzmann & Laubier, 1975 |
| status |
|
Paraonides myriamae Katzmann & Laubier, 1975 View in CoL
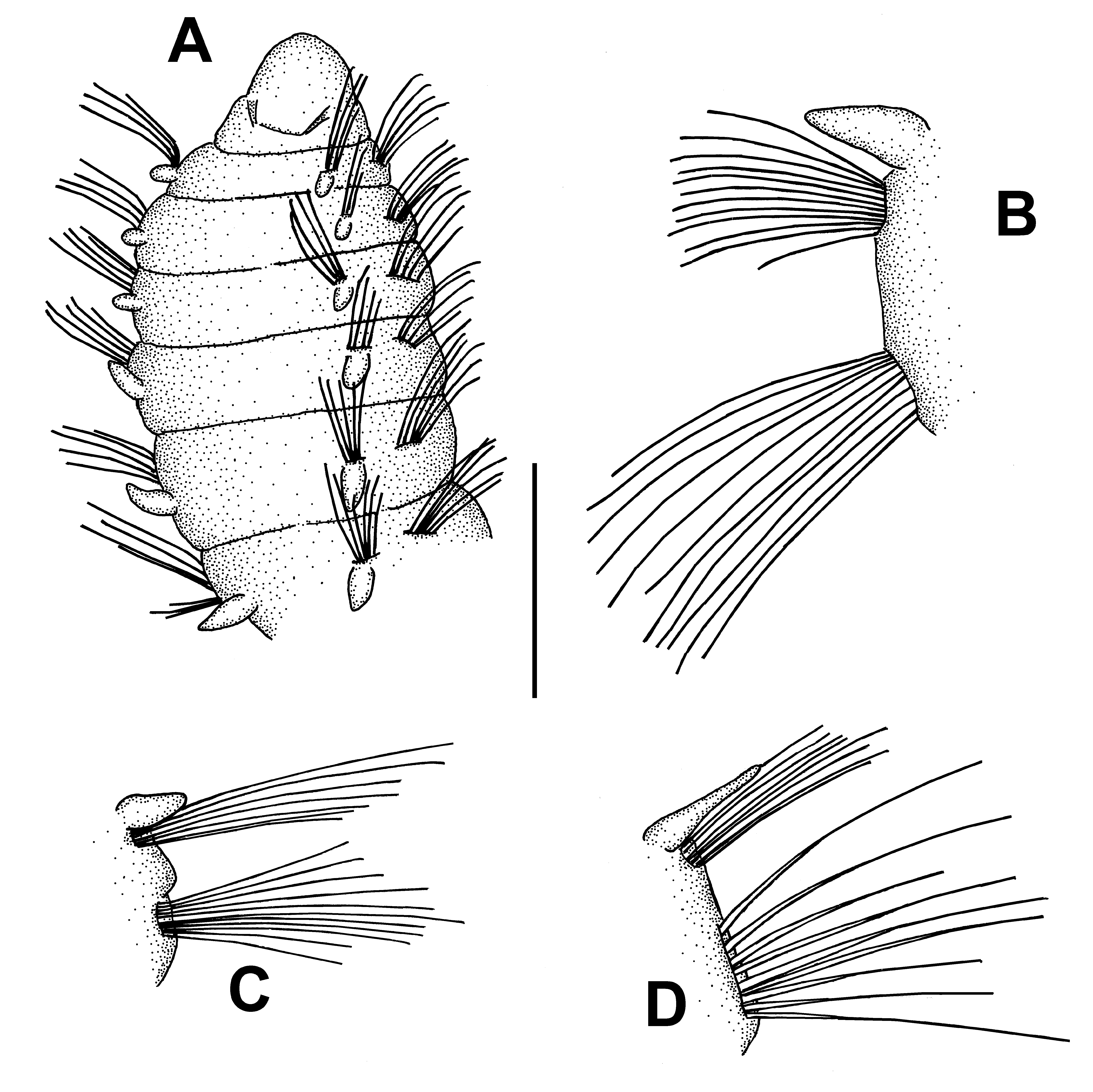
( Fig. 9 View FIGURE 9 )
Paraonides myriamae Katzmann & Laubier, 1975: 572 View in CoL –575, fig. 2; Aguirrezabalaga & Gil 2009: 640 –641, fig. 7.
Material examined. 1612 Sygna SY–09; 61.4685°N–1.9926°E; 303 m; 08.06.1999; one specimen. 1612 Sygna SY–08; 61.4509°N–1.9717° E; 290 m; 11.06.1999; one specimen.
Description. Largest specimen complete, 4.8 mm long, 0.08 mm wide for 85 chaetigers. Body slender, cylindrical, coiled when preserved. Prostomium as equilateral triangle ( Fig. 9A View FIGURE 9 ); median antenna and apical sensory organ absent; pair of nuchal organs as conspicuous oblique nuchal slits, posterior end of nuchal organs united by a transversal groove ( Fig. 9A View FIGURE 9 ). Notopodial postchaetal lobes short and piriform, similar in length along almost entire body ( Fig. 9A–C View FIGURE 9 ); those of last chaetigers digitiform and twice longer but clearly shorter than body width (Fig. D). Branchiae absent. Noto- and neurochaetae as geniculated bilimbate capillaries in first ten chaetigers ( Fig. 9A, B View FIGURE 9 ); then progressively thinner and straighter ( Fig. 9C View FIGURE 9 ); neurochaetae in posterior segments clearly thicker than corresponding notochaetae ( Fig. 9D View FIGURE 9 ); modified noto- or neurochaetae absent. Pygidium as a rounded lobe with a pair of bottle shaped anal cirri.
Remarks. Our specimens are characterized by completely lacking branchiae and modified chaetae in both rami of parapodia; because of the latter, they can be accurately referred to the genus Paraonides Cerruti, 1909 . Other diagnostic features of our specimens are the notopodial postchaetal lobes present from chaetiger 1 and similarly short along body, and the triangular prostomium bearing conspicuous nuchal organs; all of these features are recorded in both original description of P. myriamae ( Katzmann & Laubier 1975) and more recent description ( Aguirrezabalaga & Gil 2009). Only two species described within this genus lack branchiae, namely P. myriamae Katzmann & Laubier, 1975 and P. monilaris Hartman & Fauchald, 1971 . Paradoneis monilaris , from abyssal depths in central Atlantic ( Hartman & Fauchald 1971), is an altogether different species, with rounded prostomium lacking apical sensory organs and all the segments arranged in a moniliform way. Furthermore, this species lacks notopodial postchaetal lobes in chaetiger 1. Paraonella abranchiata Fauchald & Hancock, 1981 , is a species from deep waters off western coast of United States ( Fauchald & Hancock 1981) that should be assigned to Paraonides , as defined by Castelli (1988). This species bears a prostomium of similar shape to that of P. myriamae , but its nuchal organs are far less conspicuous. Moreover, P. abranchiata bears notopodial postchaetal lobes in the posterior chaetigers that are longer than the rest, similar to the body width in the corresponding segment.
Distribution. Adriatic Sea, Mediterranean ( Katzmann & Laubier 1975). Bay of Biscay ( Aguirrezabalaga & Gil, 2009). First record for Norway and adjacent waters.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Sedentaria |
|
Family |
|
|
Genus |
Paraonides myriamae Katzmann & Laubier, 1975
| Sikorski, Andrey 2017 |
Paraonides myriamae
| Aguirrezabalaga 2009: 640 |
| Katzmann 1975: 572 |