
Helvetostrea, Koppka, Jens, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3927.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:42B56D11-9B18-4FCC-B632-30A46AB0205C |
|
DOI |
https://doi.org/10.5281/zenodo.6102704 |
|
persistent identifier |
https://treatment.plazi.org/id/37F96388-804D-4814-9720-5CBAAE14C6C5 |
|
taxon LSID |
lsid:zoobank.org:act:37F96388-804D-4814-9720-5CBAAE14C6C5 |
|
treatment provided by |
Plazi |
|
scientific name |
Helvetostrea |
| status |
gen. nov. |
Helvetostrea n. gen. sequana (Thurmann & Etallon, 1862)
Figs. 14–16 View FIGURE 14 View FIGURE 15 View FIGURE 16 ; Pl. 4.7–9; Pl. 11.1–5; Pl. 12.1–2; Pl. 20.2
? 1848 Ostrea sequana —Dufrénoy & de Beaumont: p. 594.
1852a Ostrea Sequana nob.—Thurmann: p. 213 (nom. nud.).
1854 Ostrea Sequana Th. —Thurmann: p. 354 (nom. nud.). 1858 Ostrea sequana Thurm. —Oppel: p. 150, 164, 165.
1861 Ostrea Sequana, Th. —Thurmann & Etallon: p. 37, 64. 1862 Ostrea Sequana, Th. —Etallon: p. 407.
* 1862 Ostrea sequana, Th. —Thurmann & Etallon: p. 270, pl. 38, fig. 6. 1870 Ostrea Sequana, Th. —Greppin, J.-B.: p. 104.
1887 Ostrea sequana Th. —Roberts: p. 238.
1917 Ostrea Sequana (Thurm.) Etallon—Rollier : p. 589.
1971 Liostrea sequana (Thurmann, 1862) —Pugaczewska: p. 257, pl. 16, figs. 1–4.
Lectotype (designated herein). Ostrea sequana Thurmann & Etallon, 1862 , p. 270–271, pl. 39, fig. 1, left valve. The type specimen (refigured on Fig. 15.1) was not traceable in the MJSN Porrentruy and is apparently lost. However, ICZN Article 74.4 permits the designation of an illustration as lectotype when the type specimen is lost or not traceable.
Combe d'Essert-Tainie, a gorge (not traceable on the available maps of the region) but situated most likely northwest of Bressaucourt (near Chevenez, Ajoie, Canton Jura), "Marnes à Astartes" (now Bure Member, Vellerat Formation, Epipeltoceras bimammatum Zone ), Upper Oxfordian; coordinates: 47°23'17.36''N, 7°02'15.98''E; the coordinates refer to Bressaucourt.
Material. Lectotye (see above) and paralectotype (AC1766, B.4.15.4.1)—Only one small rock-sample from the type series was traceable in the MJSN Porrentruy. The small piece of grey, dense limestone contains several cross-sections of weathered and only partially preserved specimens exhibiting the typical hollow chambers of H. sequana but no other shell details. Due to the poor preservation none of the specimens is suitable to be chosen as lectotype.
The original label of the paralectotype refers to Ostrea sequana nob, Th. 1846, from Bressaucourt. However, according to Etallon’s introduction to the “Lethea bruntrutana ” ( Thurmann and Etallon, 1861, p. 3), the species name was initially a nomen nudum based on an unpublished note by Thurmann (1848) titled “Sur l’ Ostrea sequana ”. Therefore, Dufrénoy & de Beaumont (1848) and Thurmann (1852, 1854) can only have referred to this unpublished name, whereas the valid introduction is given in Thurmann & Etallon (1862).
Others—One small (“juvenile”) LV from the basal Banné Member (horizon 300, cymodoce Zone, Lower Kimmeridgian) of Vâ Tche Tchâ near Courtedoux ( VTT 001-1169b, Pl. 11.3) and more than 20 larger (“adult”) specimens (mostly LV, Pl. 4.7–8; Pl. 11; Pl. 12.1–2) from the “Virgulien” facies of the uppermost Courtedoux Member, Upper Kimmeridgian (horizons 2400–3500, mutabilis Zone, lallierianum Subzone) of Sur Combe Ronde (SCR) and Tchâfouè (TCH) near Courtedoux. The best preserved specimens are listed in the Appendix, Table 5 View TABLE 5 .
Two small and thin-shelled specimens labelled as Ostrea sequana Thurmann ( MJSN S.1337, A.2.18.2.4) from the Koby collection in the MJSN belong to an undetermined liostreine species.
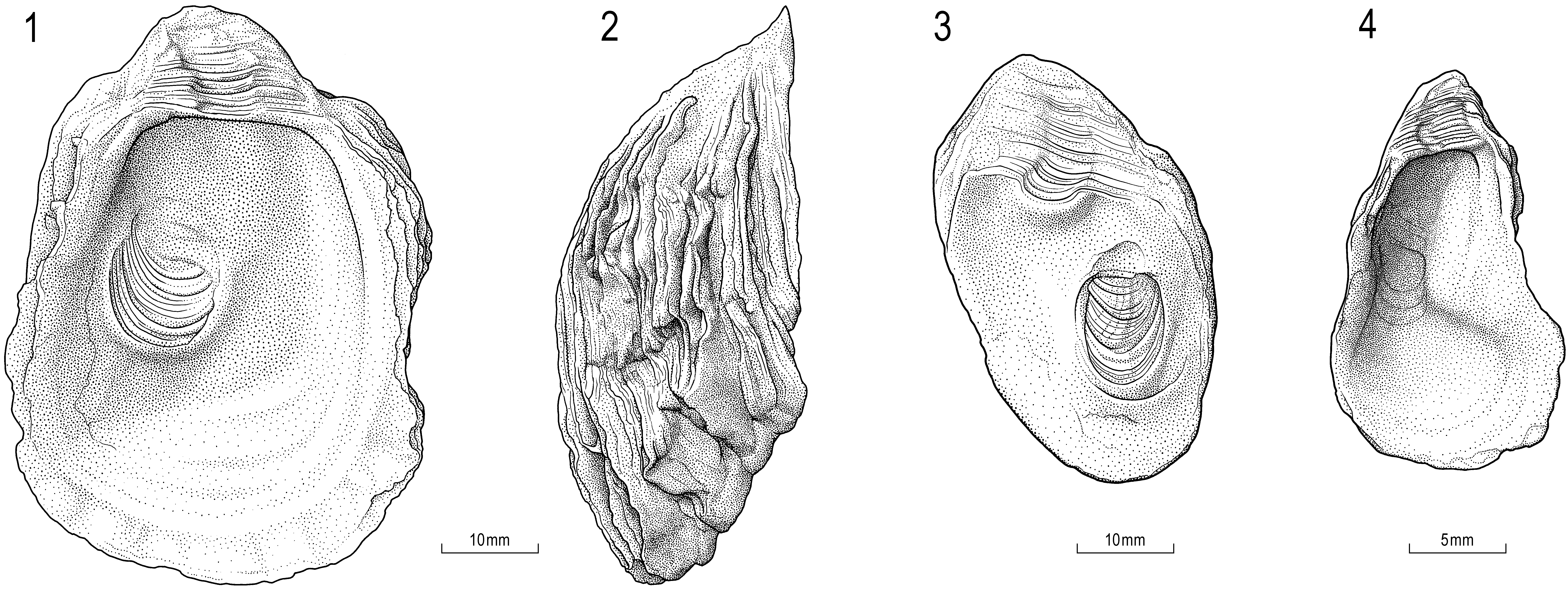
Description. Shape, size—Highly variable including subrectangular, trapeziform, subtrigonal shapes. LV sometimes much elongated in height, always higher than RV, usually moderately inflated, greatest convexity close to umbo. Attachment area commonly restricted to umbonal region, medium-sized in separately growing individuals (Pl. 4.7, 9a; Pl. 11.2, 4a) or large when growing in clusters (Pl. 4.8, Pl. 12.2a). RV (only two specimens known; Pl. 11.1b; Appendix, Table 5 View TABLE 5 ) relatively small and flat, with subrectangular shape and slightly convex, shell thickness at the ventral margin 3–4 mm. Observed maximum size 8 cm (height) (Appendix, Table 5 View TABLE 5 ).
Sculpture—LV with thick growth lamellae, irregular, imbricating (spaces between squamae filled with sediment, later overgrown by new shell foils); occasionally with few undulating radial ribs; shell surface of folds and depressions, in some cases pustular. Single RV relatively smooth, with few weak commarginal furrows and fine growth lines.
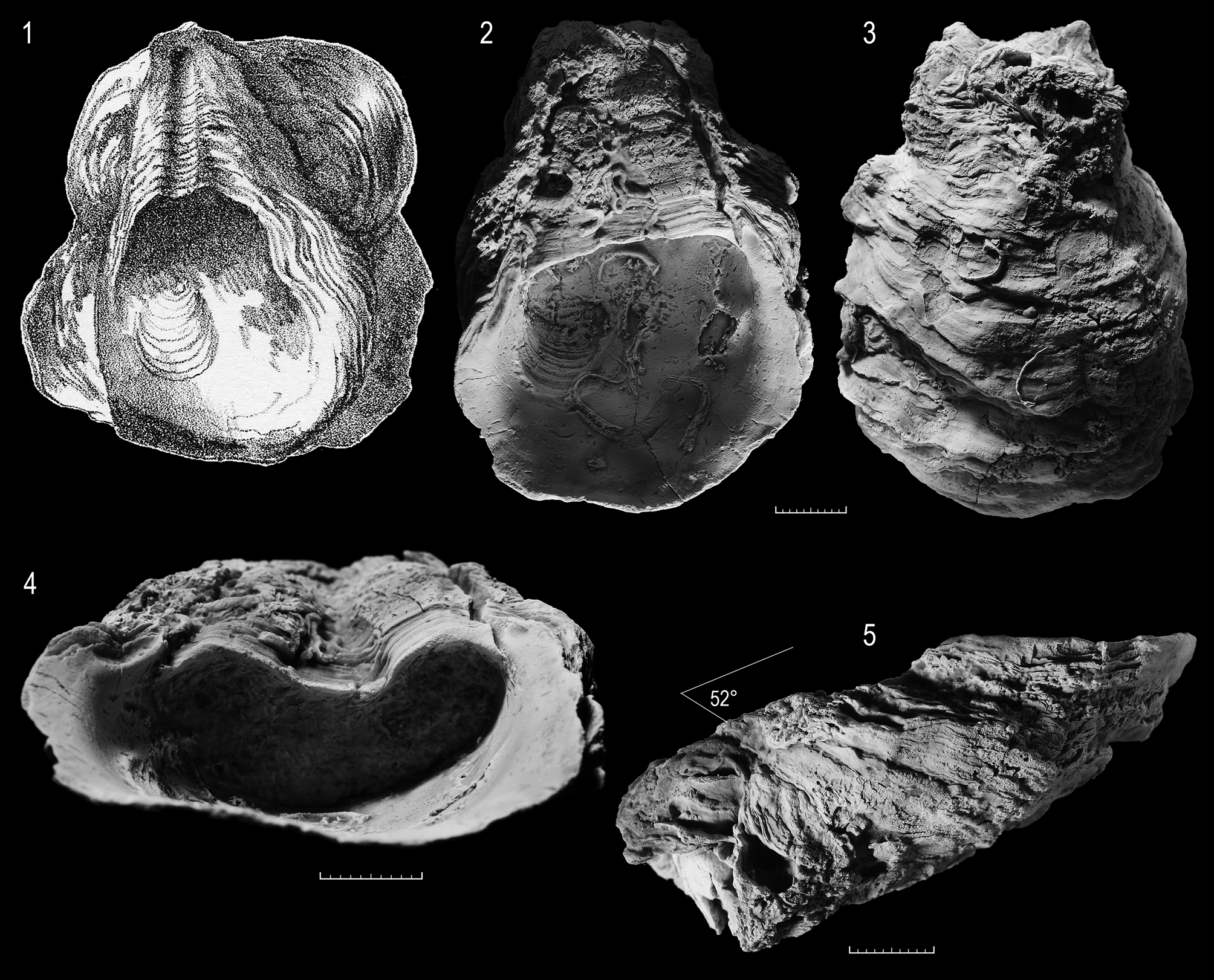
Ligament area—Overall growing ventralwards, ostreoid, with a straight or slightly undulating resilifer, rarely anteriorly directed (Fig. 14.3–4; Pl. 11.2). LV resilifer deep, flanked by equally broad, elevated bourrelets (Fig. 15.2); outer bourrelet flanks dipping steeply towards commissure (ca. 30–60°) (Pl. 11.4a; Fig. 15.5), confined anteriorly and posteriorly by deep narrow troughs (Fig. 15.2,4; Pl. 11.2). RV resilifer supported by a callous-like shell buttress (sensu Stenzel 1971, p. N1029, N973, fig. J14a,b) (Fig. 14.3; Pl. 11.1b)
Internal shell characters—LV commissural shelf, dorsally narrow and bounded by inner lateral ridge; shelf broadening ventrally. Posterior adductor scar subcircular to high-oval, dorsally truncated or weakly concave, scar slightly sunken below general depositional surface; position essentially posterodorsal (ca. 1/3 sub-posterocentral). (Fig. 14.3–4; Fig. 15.2; Pl. 11.1a,b). PAM insertion in RV with projected ventral border (Pl. 11.1b) (buttressed sensu Stenzel 1971, p. N1029). Quenstedt muscle scar not seen. Chomata and relict chomata questionable (Pl. 11.3a,b, 5c,d).
Microstructure—LV with thick regularly to partly complex cross-foliated outer-middle layer; middle-inner layers largely chambered, some (probably originally) hollow (Pl. 12.1a,b), some filled with chalky deposits (mocret); some filled with sediment or diagenetic cement (Pl. 12.2b,c; Pl. 20.2a–d).
Prodissoconch, juvenile—Prodissoconch and early juveniles unknown.
Paleoecology. At its type locality, the species formed aggregates or small buildups (“framestones”) of up to several decimetres thickness intercalated between coral-bearing, calcareous marls and unfossiliferous marls („Zone Astartienne“); the lateral extensions are unknown. Thurmann & Etallon (1861, p. 37; 1862, p. 271, pl. 39, fig. 1) found the species together with Nanogyra nana (= Exogyra bruntrutana Thurmann ), for example, in the “Lumachelle à Exogyra bruntrutana et Ostrea sequana ”. The species is associated with a diverse coral fauna of the „Zone à polypiers“ ( Thurmann & Etallon 1861, p. 37, 62), crinoids (genus Apiocrinites ) and terebratulid brachiopods.
In the study area, the species occurs as autochthonous element in horizons 2600 and 2900 –3000, and allochthonous in horizons 300 (Banné Marls) (one small juvenile LV, VTT001-1169b, Pl. 11.3; Pl. 14.6), 2400, 2700 (Pl. 11.1d), and 3500 ( Fig. 4 View FIGURE 4 C).
Horizon 2600 is a mostly fine-grained, partially iron-oolitic limestone yielding numerous aggregates of H. sequana LVs. Aggregates are reminiscent of ostreoliths described by Wilson et al. (1998), with irregularly arranged specimens of different growth stages, growing in all directions which suggests several overturns (Pl. 4.8; Pl. 12.2).
Shell surfaces are covered by serpulids and small LVs of Nanogyra nana and Gastrochaenolites borings. In one case, Lithophaga sp. is preserved in its borehole. The species co-occurs with stenohaline bivalves ( Gervillella , Cercomya, Myopholas , Anisocardia ) and the echinoid Pygurus sp.
Horizon 2900 is a fossiliferous lithoclastic limestone with numerous LVs of Helvetostrea ; co-occurring fauna includes species of Trichites , Gervillella, Cucullaea, Myopholas, Placunopsis , rare Myophorella (shells replaced by calcite), and recrystallized coral heads up to 20 cm in size. This accumulation may represent a condensation horizon. It is truncated by a hardground (horizon 3000) penetrated by Gastrochaenolites sp. and overgrown by large, strongly abraded specimens of Circunula cotyledon .
The autochthonous or parautochthonous occurrences and associated faunas suggest that Helvetostrea sequana lived preferentially in shallow, fully marine environments where it formed buildups (type locality), small clusters, or ostreoliths.
A similar setting was described by Wilson et al. (1998) for the American ostreoliths which were formed by Circunula ? strigilecula reaching sizes of up to 50 cm. They occurred on hardgrounds and oolitic shoals but also in lagoonal facies (Middle Jurassic Carmel Formation, Utah, USA). Fürsich et al. (1994) reported reefoidal frameworks several meters wide and up to 3.5 m high constructed by H. expansa , red algae ( Solenopora " portlandica "), the cyclostome bryozoan Hyporosopora portlandica (Gregory), and some cemented Plicatula damoni ( Cox, 1925) . Hoffmann & Krobicki (1989) described a buildup of 2 m lateral and 0.8 m vertical extension from the Middle Jurassic of Poland constructed by H. explanata (Goldfuss) surrounded by a dysaerobic mudstone facies. The buildup occurs in high energy sediments, deposited as an intercalation during a "regressive" phase, well documented by a shell bed with many clasts (including hiatus-concretions) with borings of Gastrochaenolites lapidicus Kelly & Bromley, 1984 and encrusting serpulids at the base of the buildup ( Hoffmann & Krobicki 1989, p. 308 – 309, fig. 3).
Comparisons. Species of Helvetostrea n. gen. and morphological similar forms are discussed below in alphabetical order.
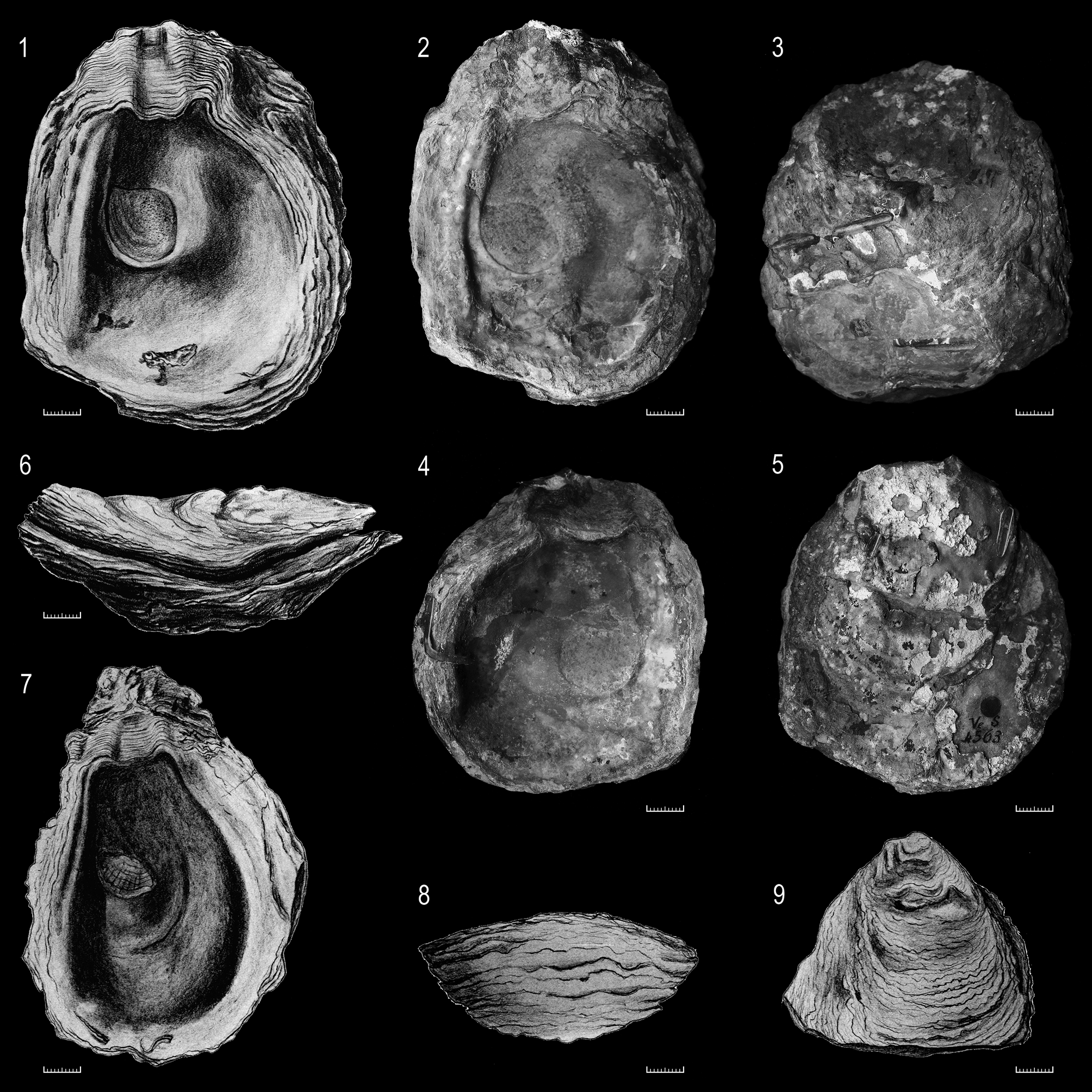
" Ostrea " caprina Rollier, 1917 (pl. 39, fig. 2, and pl. 40, fig. 2a,b; pl. 39, fig. 2 is refigured on Fig. 16.1)—The species was first mentioned by Moesch (1867: p. 148) from Gösgen near Olten (Aargau) and found in the " Perna " beds of the Geissberg Member (sensu Gygi 1969), Upper Oxfordian ( bimammatum Zone ) of northeastern Switzerland. However, Rollier (1917, p. 562) was the first to provide a valid description. The type series consisted of one bivalved specimen with both shells isolated and an unknown number of other specimens mentioned in the text from different locations, but only the figured bivalved specimen (ETHZ Ve.S. 4563) of Rollier (1917: pl. 39, fig. 2, pl. 40, fig 2a,b) is still preserved in the collections of the ETH Zürich. It is here chosen as lectotype of Helvetostrea caprina (Rollier, 1917) and figured on Fig. 16.1, 2–6.
This species is less inflated than H. sequana , more subrectangular and lacks an umbonal cavity. Shell habitus and ligament area of the lectotype LV (Fig. 16.1–2) mimic a RV, but the adductor position and presence of an attachment area confirm that it is a LV. The comparison of the original figures of Rollier (1917) with photographs of the lectotype reveals a mismatch of the real proportions and hinge morphology (Fig. 16.1–6).
" Ostrea " expansa J. Sowerby, 1819 (p. 65, pl. 238, fig. 1)—This species was first described from the Tithonian Tisbury Limestone Member of the Portland Stone Formation ( Titanites anguiformis Zone ) in Wiltshire, England. Sowerby’s figured specimen, a silicified RV, was assumed to be lost ( Cox 1929, p. 147). However, Spamer & Bogan (1989) may have found the original or a syntype and potential lectotype in the Benett-collection at the Academy of Natural Sciences of Philadelphia ( USA) which is listed as " Ostrea expansa J. Sowerby 1819 , pl. 238, top figure [1]; syntype (?), ANSP 56158 (B.C. 287; Benett's[?] no. 196); Fonthill, Wiltshire". The Fonthill Abbey Wood is situated 3 km northwest of Tisbury.
Sowerby’s figure is somewhat reminiscent of the flat, broadly crescentic shape of Deltoideum (Rollier 1917, p. 566: Deltoideum expansa ). However, the overall shape is spatulate without a concave posterodorsal margin typical of Deltoideum . Furthermore, the shell is thicker and larger, and the PAM more centrally situated. Despite these differences, Cox (1925, p. 130) assumed a transition between Deltoideum delta (Smith, 1817) [= O. deltoidea J. Sowerby, 1816 , non Lamarck 1806, = Ostrea sowerbyana Bronn, 1836 , replacement name for the former] and O. expansa . Pugaczewska (1971, p. 269), Fürsich et al. (1994) and Fürsich & Hautmann (2005) assigned the species to Liostrea . However, the spatulate shell shape and presence of large shell chambers below the ligament area suggest that O. expansa belongs to Helvetostrea . The species differs from H. sequana by a broader resilifer, slender bourrelets, a larger PAM, a convex RV and a small umbonal cavity ( Fürsich et al. 1994, p. 138, 141, fig. 5).
Remarkable is the distribution of both species in rocks representing shallow marine environments that are overlain by paleosoils with dinosaur track sites of the Lower Purbeck Formation in England ( Ensom & Delair 2007) and, respectively, the Kimmeridgian track-bearing horizons (tidal-flat laminites) of the Reuchenette Formation in Switzerland (Marty 2008).
" Ostrea " explanata Goldfuss, 1833 (b) (p. 22, pl. 80, fig. 5)—This is the oldest species that could be attributed to Helvetostrea . Goldfuss described it from a Middle Jurassic Oolite (Bajocian?) of Southern Germany (region of Bayreuth and Württemberg, type locality not specified). In Europe, it ranges from the Bajocian to the Early Callovian in England, France and Poland ( Hoffmann & Krobicki, 1989, p. 308; Pugaczewska, 1971, p. 245).
The species is usually determined as Liostrea (e.g. Pugaczewska 1971) coinciding with its ancient synonymization with Liostrea eduliformis ( Schlotheim, 1820) by Roemer (1835) and Schlippe (1888) (here considered as a Pernostrea ). Specimens figured by Goldfuss (1833)(b) show a broad but short ligament area which is also more reminiscent of Pernostrea , whereas a right valve figured by Pugaczewska (1971, pl. 9, figs. 5a,b) has a remarkably undulated hinge line and ligament similar to Helvetostrea . Hence, “ O.” explanata sensu Pugaczewska (1971) may belong to Helvetostrea . However, it differs from H. sequana by its larger size (in height), more rectangular and less inflated shape, and lack of an umbonal cavity ( Hoffmann & Krobicki, 1989, p. 308).
" Ostrea " oxfordiana Rollier, 1917 —This rare species was originally described from the "Terrain à Chailles" facies ( Pholadomya View in CoL marls, Early Oxfordian, cordatus Zone) of Klein-Kembs (Kleinkems) from the "Isteiner Klotz", a prominent ridge some kilometres north of Lörrach (Baden-Württemberg, Germany). The figured holotype (fixed by monotypy, ICZN 73.1.2.; see also ICZN 73.1.5. which implies that a holotype can consist of a set of disarticulated components) consisted of a rather well preserved LV and a corresponding but damaged RV (Rollier 1917, pl. 40, figs. 1a–c, here refigured in Fig. 16.7–9). It belonged to the Casimir Moesch collection held at the ETH in Zürich (Ve. S. 5146, according to Rollier 1917); unfortunately, its current whereabouts is unknown (Pika- Biolzi, pers. comm. 2011).
The species could be a Helvetostrea that differs from H. sequana by having a more pronounced umbo, possibly due to a smaller attachment area, shorter ligament area, lesser convexity of the LV, more prolonged commissural shelf, and a shallower umbonal cavity.
In contrast, Liostrea oxfordiana sensu Pugaczewska (1971, p. 249, pl. 13, figs. 5 – 9) from the Lower Kimmeridgian of West Pomerania ( Poland) as well as Liostrea quadrangularis Arkell, 1927 , sensu Pugaczewska (1971, p. 250, pl. 8, figs. 1–4) are more likely phenotypes of Liostrea polymorpha (Münster in Goldfuss, 1833)(b) ( Pugaczewska 1971, pl. 19, figs. 5 – 8). All these species differ from H. sequana by a markedly opisthogyrate ligament area, and a weakly sickle-shaped outline .
" Ostrea " ermontiana Etallon, 1862 (in Thurmann & Etallon, 1861 –1864, p. 270, pl. 38, fig. 6)—The species was erected as a substitute for Ostrea gryphoides Contejean, 1859 , which is a junior homonym of Ostrea gryphoides von Zieten, 1830, and Ostrea gryphoides Schlotheim, 1813 . Contejean’s (1859, p. 320, pl. 25, figs. 1–5) material came from the Upper Oxfordian “Calcaires à Cardium ” and Lower Kimmeridgian “Marnes et Calcaires à ptérocères” of Montbéliard ( France) (housed at the Musée Cuvier in Montbéliard, MC 27E125 = LV in Contejean 1859, pl. 25, figs. 1, 3; MC 27E126 = LV, pl. 25, figs. 2, 4; MC27E127 = LV, pl. 25, fig. 5). However, all specimens are exfoliated internal moulds with few shell remains. Because the specimens are, at face value, indeterminable, and because there is no other material mentioned, the figured LV of O. ermontiana is here considered as lectotype. The specimen was found in the "Hypostrombien inférieur" (= Nautilid Beds, Reuchenette Formation, Lower Kimmeridgian) of Fahy near Porrentruy, but neither the holotype nor any other material potentially representing this species was found in the Jurassica-Museum (MJSN) Porrentruy. One additional complete and well-preserved specimen, determined as Ostrea ermontiana Etallon , was found in the collection Scheurer (MC-27E122) from the “Calcaire à Corbis ” (cymodoce Zone) at Baume in Audincourt ( France). It also contains a note by Scheurer that O. gryphoides is O. ermontiana .
The figured lectotype in Thurmann & Etallon (1862) is a small LV gryphaeoid shell (L 5.5 cm, H 3.5 cm) with an orthogyrate ligament area, which appears as drawing similar to H. sequana . After comparison of the available material an assignment of O. ermontiana to Helvetostrea can be excluded and the species can certainly be assigned to Gryphaea (Bilobissa) .
Differences to H. sequana are the presence of a deep radial posterior sulcus (Thurmann & Etallon 1862, pl. 38, fig. 6, posterior view), which is characteristic of Bilobissa, the thin and not chambered shell, the nearly smooth LV and a RV with antimarginal threads (preserved on specimen MC-27E122). The relationship of Gryphaea (Bilobissa) ermontiana (Etallon in Thurmann & Etallon, 1862) to Gryphaea (B.) dilatata (J. Sowerby, 1816) has not been investigated, but the occurrence of this species in the Early Kimmeridgian (cymodoce Zones) of the Reuchenette Formation in Switzerland and nearby France represents one of the last records of Gryphaea in Europe, beside its record from Poland ( Pugaczewska 1971, p. 277).
“ Crassostrea ” tetoriensis Komatsu et al., 2002 —This oyster is only known from the Middle Jurassic of Japan (Bathonian, Ushimaru Formation, Tetori Group) (see Fujita 2003, fig. 2, for stratigraphy). Like Helvetostrea species, “ C.” tetoriensis Komatsu et al. 2002 , p. 1043, figs. 3, 5–7 developed numerous chambers in the LV, a deep umbonal cavity, an elongated ligament area, lack of chomata, and inequivalve shells without radial ornament. It also occurs below non-marine deposits with dinosaur bones and tracks (compare paleoecology of H. sequana ). The species differs from Helvetostrea sequana by its very high, elongate-spatulate outline ( C. tetoriensis : H/L ratio 2.9–3.7; H. sequana H/L 1.23–1.86), thin, mainly simple compact foliated shells with small chambers, usually filled with chalky deposits, a small reniform PAM not supported by a ventral shell swelling (buttress), narrow bourrelets and tiny attachment area ( Komatsu et al. 2002, p. 1043). In addition, the species was most probably adapted to euryhaline, muddy, intertidal, soft bottom environments.
“ Ostrea ” hebridica ( Forbes, 1851) — The species was originally described from the upper part of the Middle Jurassic (Bathonian) “Great Estuarine Series” of Loch Staffin, Isle of Skye, Inner Hebrides of Scotland ( Arkell 1934, p. 11–12).
Most of the previous authors, including Hudson & Palmer (1976, p. 87), accepted a close relationship between “ O.” hebridica and Praeexogyra acuminata . However, “ O.” hebridica is extensively chambered and euryhaline ( Hudson & Palmer 1976) whereas P. acuminata is an essentially compact foliated ( Siewert, 1972) euhaline species, co-occurring with ammonites in England as well as in Switzerland ( Arkell 1934; own data from the “ Acuminata Beds” of the Hauptrogenstein Formation, Germany and Switzerland).
Shell chambering and euryhaline preference may indicate a closer, perhaps congeneric relationship with “ C. ” tetoriensis . Shell chambering could also indicate a link to Helvetostrea . However, the species differs from all Helvetostrea species by its more ventrally positioned, crescentic to reniform PAM without ventral shell buttress, with a pointed umbo, usually tiny attachment area, short ligament area, generally slender outline , curved posterodorsal margin, and antimarginal riblets and furrows, especially the early ontogenetic ribbed “ Catinula View in CoL ” stage ( Arkell 1934; Hudson & Palmer 1976). In addition, all Helvetostrea species are euhaline.

| VTT |
VTT Biotechnology, Culture Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Helvetostrea
| Koppka, Jens 2015 |
tetoriensis
| Komatsu et al. 2002 |
tetoriensis
| Komatsu et al. 2002 |
explanata sensu
| Pugaczewska 1971 |
Liostrea oxfordiana sensu
| Pugaczewska 1971 |
Liostrea quadrangularis
| Arkell 1927 |
caprina
| Rollier 1917 |
Helvetostrea caprina
| Rollier 1917 |
oxfordiana
| Rollier 1917 |
ermontiana
| Etallon 1862 |
Ostrea gryphoides
| Contejean 1859 |
hebridica (
| Forbes 1851 |
Ostrea sowerbyana
| Bronn 1836 |
explanata
| Goldfuss 1833 |
Liostrea eduliformis (
| Schlotheim 1820 |
expansa
| J. Sowerby 1819 |
Ostrea expansa
| J. Sowerby 1819 |
Deltoideum delta
| Smith 1817 |
O . deltoidea
| J. Sowerby 1816 |
Gryphaea (B.) dilatata
| J. Sowerby 1816 |
Ostrea gryphoides
| Schlotheim 1813 |