
Oligosoma pachysomaticum ( Robb, 1975 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4688.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:DDA1FDD7-895E-4667-ACA7-4003B45DFF5D |
|
persistent identifier |
https://treatment.plazi.org/id/04197D19-FFCD-FF24-FF59-FB2E62F9F2EA |
|
treatment provided by |
Plazi |
|
scientific name |
Oligosoma pachysomaticum ( Robb, 1975 ) |
| status |
|
Oligosoma pachysomaticum ( Robb, 1975)
Figure 3 View FIGURE 3
Synonomy
Leiolopisma pachysomaticum Robb, 1975
Cyclodina oliveri (in part) Hardy, 1977
Cyclodina pachysomaticum (in part?) Wells & Wellington, 1985
Cyclodina oliveri (in part) Patterson & Daugherty, 1997
Cyclodina oliveri (in part) Chapple et al. 2008b
Oligosoma oliveri (in part) Chapple et al. 2009
Holotype: AM H538 (author’s number R67), adult female, Ruamahua-nui Island , Aldermans group, November 1972, A. H. Whitaker.
Diagnosis. An Oligosoma with a squarish mid-body cross-section; opaque, divided palpebral disc; a prominent tear-drop mark beneath each eye; iris light brown to orange; mid-body scale rows 32–36; ventral scale rows 72–88; eye diameter> distance from eye to mouth; adult SVL ≤ 84 mm; dorsal markings dominated by fine black flecking; belly cream and marked with numerous individual fine dark flecks. Distinguished from similar congeners (which share both the squarish body cross-section and the tear-drop subocular marking) as follows: alani (sympatric) has a black iris, prominent dorsal blotches and is much larger at up to 142 mm SVL; macgregori has longitudinal streaks on the dorsum and a uniform belly; ornatum (potentially sympatric) is smaller (SVL usually <80 mm), usually has a yellow belly and usually has a tapering pale dorso-lateral stripe; oliveri has 36–44 mid-body scale rows, 88–97 ventral scale rows, dorsal and ventral patterns dominated by pale blotches, and a smaller eye (diameter <distance from eye to mouth); roimata is smaller at up to 65 mm SVL and has only 1 primary temporal scale (versus 2 in pachysomaticum ); townsi usually has only 1 primary temporal scale and has few ventral markings; and whitakeri (sympatric) is larger at up to 101 mm SVL, has a dark-brown iris and a yellow-orange venter.
Etymology. From the Greek pachys (thick) and soma (body). In the absence of an established vernacular name for Oligosoma pachysomaticum I suggest ‘Coromandel skink’ to reflect the general location of the surviving populations.
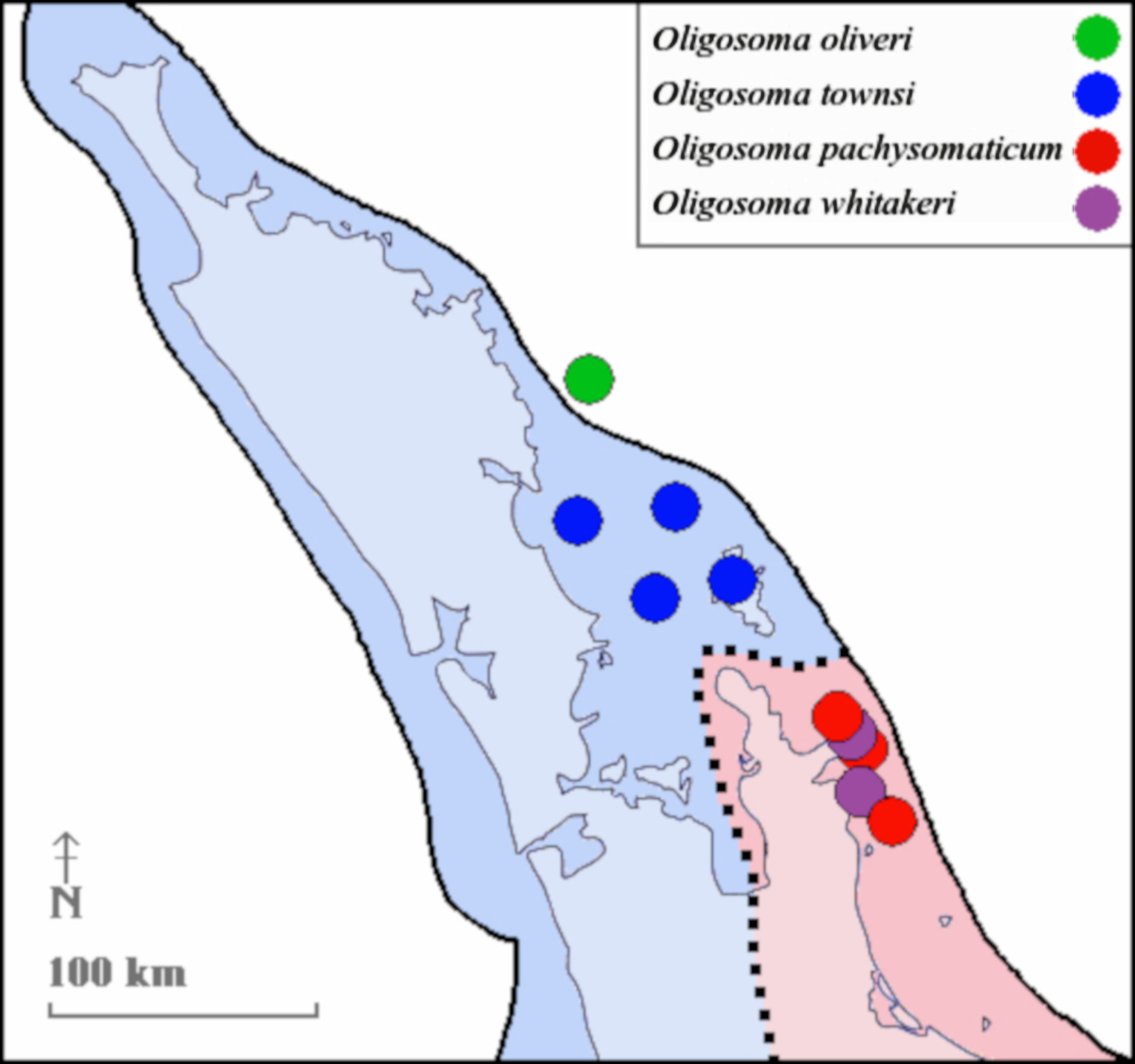
Distribution. Known from a total of eight islands in the Mercury Group (Middle, Green and Korapuki Is.), Alderman Group (Half, Hongiora, Ruamahuanui and Ruamahuati Is.) and the Ohinau Group (Old Man Rock), all off the east coast of the Coromandel Peninsula ( Figure 1 View FIGURE 1 ; Table 1 View TABLE 1 ; Chapple et al. 2008b).
Ecology. The following information is based on a study of the species on Green Island in the Mercury group by Southey (1985). Lives primarily among deep leaf litter on the floor of coastal forest and scrub, being most abundant among regenerating scrub where the leaf litter is deepest. Nocturnal with peak activity in the first few hours of darkness, and only rarely sunbasks or forages by day. Co-exists in this habitat with two much larger skinks, Oligosoma alani (≤ 142 mm SVL) and O. whitakeri (≤ 101 mm SVL) and is significantly less abundant than these, i.e. O. pachysomaticum number 46–91/ha, O. alani 277–779/ha and O. whitakeri 161–903/ha. Also co-exists with the smaller forest/coastal scrub species O. aeneum (Girard) (76 mm). Between 2–4 young are produced in April. A wide variety of invertebrates are preyed on as well as some fruit. Predators include centipedes and tuatara.
Biogeography. The islands on which Oligosoma pachysomaticum occurs were connected to the New Zealand mainland during the Otira glacial maximum 18–20,000 years B.P., when sea-levels fell to 110 m below present levels (Haywood 1991; Kirkpatrick 1999). As such, O. pachysomaticum will likely have been present on the mainland until displaced by the introduction of predatory mammals by people, a process which began between 700–800 years ago ( Wilmshurst et al. 2008). Subfossil remains attributable to the O. oliveri species-complex (but not O. whitakeri , which is distinguishable) have been found on the mainland in the nearby Bay of Plenty area ( Chapple et al. 2008b); based on proximity to extant populations these were most likely O. pachysomaticum .
Phylogeny. The two sampled island groups of Oligosoma pachysomaticum (i.e. Alderman and Mercury) exhibit a similar level of divergence among mitochondrial DNA sequences as they each do to O. oliveri , which could indicate a split between 1.07–1.43 million years ago ( Chapple et al. 2008a, b). This is much earlier than the actual (<20,000 year) split between their islands and as such could point to cryptic speciation within O. pachysomaticum as it is here defined. However, there is yet no corroboration for such a timeframe and if anything, the biogeographic and morphological evidence must cast doubt on this interpretation. Alternative explanations to be considered including a mitochondrial split much older than the actual split of the skink populations, an accelerated rate of genetic drift in one or other population, and genetic introgression from an unknown and probably now extinct lineage. For this reason the age of the divergence between the Alderman and Mercury populations is best regarded as unresolved.
Recognition. Oligosoma pachysomaticum is here reinstated as a species separate from O. oliveri because multiple lines of evidence suggest a geographical isolation that has precluded interaction for at least 1–2 million years (see ‘Biogeography’ above and ‘Phylogeny’ for O. oliveri ), in combination with a distinct niche (as defined by their respective sympatric competitors, see ‘Ecology’ for each species, above), and also morphological differentiation ( Table 2 View TABLE 2 ) which demonstrates that during this isolation the two lineages have been evolving, to a significant degree, along separate evolutionary trajectories. O. pachysomaticum meets a number of the species definitions (e.g. ‘Simpsons’, ‘niche’, ‘phylogenetic III’, ‘phenetic’, ‘genotypic’) within the encompassing ‘general-lineage’ speciesconcept of de Queiroz (1998, 2007).
Remarks. Hardy’s (1977) decision to place Oligosoma pachysomaticum in the synonymy of O. oliveri was justified by the discovery of populations that appeared to be intermediate in phylogeny, morphology and biogeography. However, the decision by Chapple et al. (2008b) to retain them as a singular entity, after removing all of the intermediate populations again as O. townsi , was less convincing because the phenotypic and biogeographic evidence now showed a substantial differentiation between the two. O. pachysomaticum is here returned to its original concept, i.e. that of Robb (1975), who provides a full morphological description.

| AM |
Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.