
Oeneis tanana A. Warren & Nakahara
|
publication ID |
https://doi.org/10.5281/zenodo.270001 |
|
DOI |
https://doi.org/10.5281/zenodo.6069227 |
|
persistent identifier |
https://treatment.plazi.org/id/03BB87A7-FFD3-FFFD-8AB1-FEC4FACCAF5F |
|
treatment provided by |
Plazi |
|
scientific name |
Oeneis tanana A. Warren & Nakahara |
| status |
sp. nov. |
Oeneis tanana A. Warren & Nakahara , sp. nov.
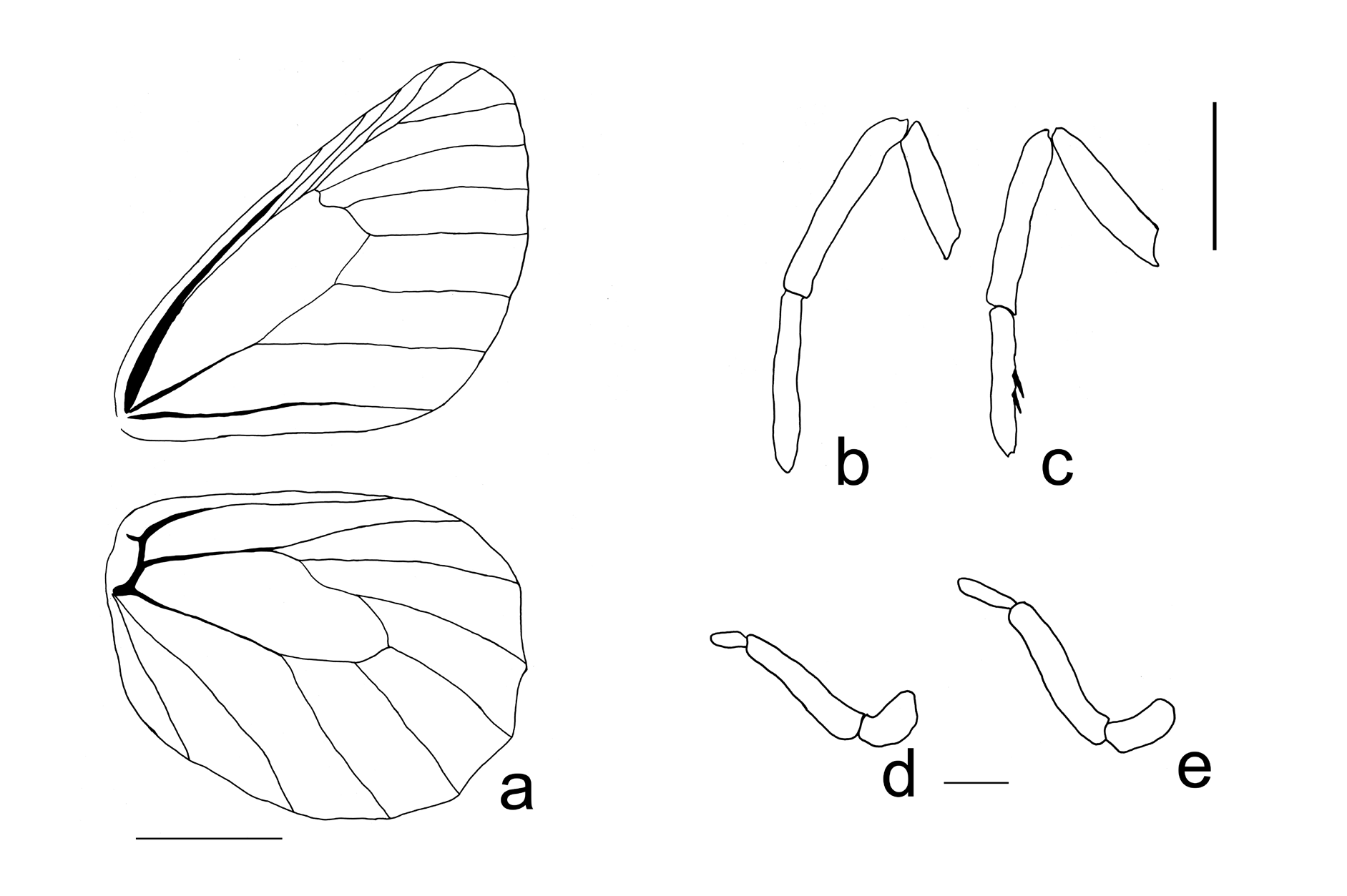
( Figs. 1 View Figure 1 , 3-4 View Figure 3 View Figure 4 , 6 View Figure 6 a-c)
Zoobank LSID: urn:lsid:zoobank.org:act:AC40896F-1D0B-4090- A52F-94EBD739D62F
M A LE. Head: Eyes brownish, naked; labial palpi ( Figs.3 View Figure 3 d,e) first segment short, covered with long dark-brown hair-like modified scales ventrally, 3-4 times as long as segment width, white scales laterally, longer white hair-like scales dorsally; second segment similar to first in scale orientation,about three times longer than first segment; third segment similar to first and second segments in scale orientation, shorter than first segment in male, same length in female; antennae approximately two-fifths length of forewing costa, 40 segments (n=1), pedicel about half as long as scape, with distal 15-16 segments comprising club. Thorax: Dorsally black, covered with golden hairlike modified scales; ventrally black, golden hair-like modified scales sparse. Legs ( Figs. 3 View Figure 3 b,c): Foreleg tarsus slightly longer than tibia, femur slightly shorter than tibia; midleg and hindleg similar in length; femur black, adorned with long dark-brown hair-like modified scales ventrally, greyish scales scattered dorsally; tarsus and tibia of midleg and hindleg covered with greyish scales,dark brown hair-like modified scales present on distal half of tibia, tibia and tarsus adorned with spines, pair of relatively short tibial spurs located at ventral side of distal end of tibia. Abdomen: Eighth tergite elongated, approximately 1.5 times longer than seventh tergite, dorsal surface apparently weakly sclerotized; eighth sternite small, approximately two-thirds length of seventh sternite, apparently uniformly sclerotized.
Genitalia ( Figs. 4 View Figure 4 a-e): Tegumen shaped somewhat like a ‘megaphone’ in lateral view, dorsal margin of tegumen slightly concave; uncus tapered towards end, slightly curved in lateral view, curved posterior end of uncus rounded in lateral view, slightly longer than dorsal margin of tegumen in lateral view, dorsally seteous; brachia almost pararell to uncus in dorsal view, apex slightly hooked, roughly half length of uncus; ventral arms of tegumen partially fused to anterior margin of tegumen, thus form of anterior edge of tegumen somewhat like a plate in dorsal or posterior view; appendix angularis present; saccus relatively short, similar in length to brachia, dorsal arms of saccus combined with ventral arms of tegumen; juxta present; valva with scattered setae, positioned at approximately 30º angle to horizontal, distal half of valva roughly trapezoidal in lateral view with angular apex, ‘tooth’ present at middle section of dorsal margin of valva in lateral view, middle section of ventral margin of valva convex in lateral view, basal one third of dorsal margin concave; aedeagus similar in length to tegumen plus uncus, almost straight in lateral view, adorned with a variable number of short spins, open anterodorsally.
Wing venation and shape ( Fig. 3 View Figure 3 a): Mean forewing length = 26.7 mm (n = 20). Forewing recurrent vein absent; basal swelling of forewing cubital vein absent; hindwing humeral vein developed;shape typical of other members of the O. chryxus complex. Wing pattern ( Figs. 1 View Figure 1 a-l): Dorsal forewing ground color dark brown; androconial dark scales approximately 1mm in width, present at distal end of discal cell along cubital vein, base of cells M3, Cu1 and Cu2; color and density of androconial scales highly variable; black submarginal ocellus in cell M1 often with indistinct creamy pupil in center; submarginal ocelli variably present in cells M3, Cu1, with or without pale pupils; submargin and margin of forewing variably overscaled with reddish or pale ochre, sparse or absent over and adjacent to wing veins, creating a series of irregularly-shaped patches separated by dark wing veins; fringe scales white and greyish. Dorsal hindwing ground colour same as forewing, with variable intensity of reddish or pale ochre overscaling; black ocellus in cell Cu1 variable in development, from bold to absent, often with indistinct creamy pupil in center; fringe scales white and greyish. Ventral forewing ground colour greyish-ochre; costal region (area basal to subcostal vein) mosaic of black and white, extending to apex, then along margin to cell R5, and variably to cells M2 or M3; numerous dark brown fragmented markings in discal cell,dark brown streak along M2-M3; dark brown undulating band extending from costa, distal to discal cell, fading distally in cell M2, curved inwards below M3 and extending to cell Cu2; black ocellus in cell M1 generally with creamy pupil in center; ocelli in cells M3 and Cu1 variably present, smallest in M3, with or without pale pupil; outer margin of forewing darker; fringe as described for upperside. Ventral hindwing ground colour indiscernible; wing veins highlighted with a variable amount of whitish scaling; costal region (area above subcostal vein) mosaic of black and white, extending along length of costa; pattern elements as follows, from base to distal margin: basal area mosaic of dark brown irregular markings with dark ochre background,followed by a whitish area with sparse dark brown irregular markings; dark brown sinuate band extending from costa to outer margin, approximately 1mm in width, roughly traversing in an outward direction until cubital vein, then roughly inward below cubital vein; area distal to this band mosaic of dark brown irregular fragmented markings with dark ochre and/ or greyish white ground colour; second dark brown sinuate band extending from costa to outer margin, similar in width to previous band, roughly traversing in outward direction until origin of M3, then roughly inward below this point; area distal to this band broadly white, wider than previous band; area distal to this (submargin and margin) mosaic of dark brown irregular fragmented markings with dark ochre and/or greyish ground color, darkest along margin; trace of pale submarginal ocelli variably present in cells Rs, M1, M 2 and M 3, black ocellus in cell Cu1 variably present, often with creamy pupil in center; fringe as described above.
FEMALE. Similar to male, except as follows: foretarsus not segmented although adorned with spines; mean forewing length = 26.9 mm (n = 10); wing shape rounder and broader, lacking forewing androconia and surrounding darkened area. Genitalia ( Figs. 4 View Figure 4 f-h): Lamella antevaginalis well developed, vertical projection under ostium bursae present and sclerotized, anterior portion of lamella antevaginallis forming a plate below this vertical projection; weakly sclerotised ventral region present in seventh and eighth intersegmental membrane, apparently fused with anterior portion of lamella antevaginalis; most of ductus bursae sclerotised; ductus seminalis located at base (posterior end) of corpus bursae; corpus bursae roughly oval, extending to third abdominal segment; two brown signa located at ventral side of corpus bursae, signa prominent and parallel to each other, spines of signa developed.
COI ‘Barcode’ sequence: vouchers CCDB-0 5786 D0 8, KWP:Ento:29760, NVG-5202, NVG-5203, 658 base pairs:
A A C T T T A T A T T T T A T T T T A G G A A T T T G A G C A G G T A T A G T A G G A A C A T C T C T T A G T C T T A T T A T T C G A A C A G A A T T A G G T A A C C C A G G A T C T T T A A T T G G A A AT G A C C A A AT T TATA ATA C TAT T G T TA C A G C T C A T G C T T T T A T T A T A A T T T T T T T T A T A G T T A T A C C A A T T A T A A T T G G G G G A T T T G G A A A T T G A C T A A T T C C T C T A A T A C T T G G A G C C C C T G A T A T A G C C T T C C C C C G A A T A A A T A A T A T A A G A T T T T G A C T T T T A C C C C C T T C T T T G A T A C T T T T A A T T T C A A G C A A T A T T G T T G A A A AT G G A G TA G G A A C A G G AT G A A C A AT T T A C C C C C C T C T C T C A T C T A A T A T T G C C C A T A G A G G A T C T T C T G T T G A T T T A G C A A T T T T T T C T T T A C A T T T A G C T G G A A T T T C T T C T A T T T T A G G A G C T A T T A A T T T T A T T A C A A C A A T T A T T A A T A T A C G A A T T A A T A A T A T A A C T T A T G A T C A A A T A C C T T T A T T T G T T T G A G C T G T A G G A A T T A C A G C T T T A T T A T T A T T A C T C T C T C T T C C T G T A T T A G C T G G G G C A A T C A C T A T A C T T C T T A C A G A T C G A A A T T T A A A T A C T T C A T T T T T T G A T C C A G C A G G A G G G G G A G A C C C T A T T T T A T A T C A A C A T T T A T T T
Types. Holotype male ( Fig. 1 View Figure 1 c) with the following labels: white, printed: AK: TANANA VALLEY / 5 MI. S. OF TOK, TOK / CUT- OFF AT BUTCH / KUTH RD. VI-17-18-99 / LEG. M.G.Douglas /; white printed: J. D. Turner ex / Malcolm Douglas / colln. / MGCL Accession # 2009-26 /; red, printed: HOLOTYPE / Oeneis tanana / A. Warren & Nakahara /. The holotype is deposited in the McGuire Center for Lepidoptera and Biodiversity, Florida Museum of Natural History, University of Florida (MGCL). Paratypes (326♂, 79♀) from: USA: ALASKA: Alaska Hwy., mi. 1270, 2000’, 11-VI-1999, J. L. Harry (9♂, 1♀ MGCL); Alaska Hwy. (Hwy. 2), mi. 1289.55, 63°13,9’N 142°17.9’W, 1800’, 15-VI-1997, C. D. Ferris (13♂, 1♀ CDF); Alaska Hwy., mi. 1289.55, Midway Lake, gravel flats on hillside above road, 15 -V I-1997, K. W. Philip (7♂, 2♀ KWP; UAM100190535-UAM100190543); Alaska Hwy., mi. 1316, 20-VI- 1955, J. & F.Preston (1♂ MGCL); Alaska Hwy.(Hwy.2), mi. 1354.2, 1800’, 63°35’N 143°55’W, 15-VI-1995, C. D. Ferris (2♀ CDF); Alaska Hw y., mi. 1371, 28 -V I-1970 (1♀ MGCL); Alaska Hwy. (Hwy. 2), mi. 1410, 1250’, 61°56.7’N 145°23.7’W, 15-VI-1995, C. D. Ferris (1♂, 1♀ CDF); Alaska Hwy., mi. 1410, Spruce Road, powerline cut in taiga, grass and flowers, 1240’, 17-VI-1997, K. W. Philip (2♂ KWP; UAM100060877, UAM100060878); Alaska Hwy., mi. 1410, 12 mi. SE Delta Jct., 1200’, 15 -V I-2001, J. L. Harr y (1♀ MGCL); Anderson, 1 mi. E, 500’, 4-VI-1999, J. L. Harry (1♂, MGCL); Hwy. 1, 5 mi. S Tok, 1700’, 10-VI-1999, J. L. Harry (18♂, 2♀ MGCL); 12-VI-1999, J. L. Harry (21♂ MGCL); 13-VI-1999, J. L. Harry (13♂ MGCL); 17-VI-1999, J. L. Harry (26♂, 6♀ MGCL); 18-VI-1999, J. L. Harry (1♂, 1♀ MGCL); Hwy. 1, 5 mi. S of Tok, 1700’, 63°16’N 143°02’W, 13 -V I-1995, C. D. Ferris (2♂, 1♀ MGCL SN-15 -145 -♂, SN-15-155-♂, SN-15-149-♀); 14 -V I-1995, C. D. Ferris (8♂ MGCL; incl. SN-15-147, SN-15-154, SN-15-156, SN-15-157); 14-15-VI-1995, C. D. Ferris (46♂, 17♀ CDF); 6-VII-1995 C. D. Ferris (1♀ MGCL, SN-15-151); Hwy. 1, 5.0 ± 0.5 mi. S of Tok, [1700’], 63°16.04’N 143°01.9’W, 5-VI-1997, C. D. Ferris (1♂, 1♀ CDF); 14-VI-1997, C. D. Ferris (52♂, 21♀ CDF); 1-VII-1997, C. D. Ferris (3♂, 4♀ CDF); Nenana, 400’ [351’], 4-VI- 1999, J. L. Harry (6♂ MGCL); 6-VI-1999, J. L. Harry (1♂ MGCL); Northway Airport, 1700’, 11-VI-1999, J. L. Harry (1♂ MGCL); Northway Airport, 7 mi. off Alaska Hwy., flower-filled lawns and fields, 1700’, 15-VI-1997, K. W. Philip (1♀ KWP UAM100190552); Old Alaska Hwy., 3 mi. NE Tok, 1600’, 12-VI-1999, J. L. Harry (2♂ MGCL); Richardson Hwy., mi. 229 [vic. Black Rapids], [2083’], 26-VI-1971, C. D. Ferris (1♀ CDF); Tanana River, 21 mi. SW Fairbanks, 400’ [Bonanza Creek Experimental Forest], 18-V-1997, J. L. Harry (1♀ MGCL); Tanana Valley, 5 mi. S of Tok, Tok Cutoff at Butch Kuth Rd., 17-18-VI-1999, M. Douglas (53♂, 7♀ MGCL); Tok, 17-VI-1971, L. Jennings (1♂ KWP; UAM100379347); 9-VI- 2005, Szymczyk (1♂ JPB); Tok Cutoff, 5 mi. S of Tok, Butch Kuth Ave., roadside flowers in open aspen/spruce forest, 13-VI-1995, K. W. Philip (11♂, 3♀ KWP; UAM100379326-UAM100379329, U A M 10 0 3 7 9 3 4 4 - U A M 10 0 3 7 9 3 4 6, U A M 10 0 3 7 9 3 6 7 - UAM100379369, UAM100379384-UAM100379387); 14-VI-1995, K. W. Philip (25♂, 3♀ KWP; UAM100379330-UAM100379343, UAM100379370-UAM100379383).
Additional material examined
Oeneis tanana : “nr. Nome, Alaska”, no date, no collector indicated (1♀ MGCL).This specimen was not included in the type series, since it is the only known specimen of O. tanana labeled from outside the Tanana River drainage, and it lacks the collection date and name of the collector; we suspect it is mislabeled. Considerable collecting efforts have been made in the Nome area, yet no material of O. tanana has been reported.
Oeneis chryxus : CANADA: BRITISH COLUMBIA (74♂, 29♀): Alaska Hwy. km. 600, 11-VII-1984, J. & F. Preston (1♂ MGCL); Alaska Hwy., mi. 392, mountain S of Summit Pass, 4000-7000’, 22-VII-1948, W. Hovanitz (1♀ MGCL); Alaska Hwy., mi. 400, Summit Lake, 4200-5000’, W. Hovanitz (2♂, 2♀ MGCL); Alaska Hwy., mi. 409, McDonald Ck., 6-VII-1948, W. Hovanitz (2♂ MGCL); Alaska Hwy. mi. 415, Racing R., 6-VII-1948, W. Hovanitz (4♀ MGCL); Alaska Hwy., MP 417, 18-VI-1970, A. O. Detmar (1♂ MGCL); Atlin, 600-900m, 22-VI-1991, J. Reichel (1♂, 1♀ MGCL); Atlin, 800m, 26-VI-1991 (1♂ MGCL); 23-VI-1991 (1♀ MGCL); Atlin Rd., 3 mi. N of Atlin, 2300’, 30-VI-1985, C. D. Ferris (4♂, CDF); Atlin Rd., 3 mi. N of Atlin to Snafu Creek, 30-VI-1985, C. D. Ferris (1♂ CDF); Coalmount, 5-VII-1968, S. Shigematsu (4♂ MGCL); Crater Mtn., W of Keremos, 1-VII-1981, C. D. Ferris (2♂ CDF); Creston, Thompson Pk., 15-VII-1976 (3♂ MGCL); Gibson Pass, Manning Park, 5000’, 16-VII-1979, C. Guppy (2♂ MGCL); 23-VII-1983 (1♂ MGCL); Haines Hwy., mi. 78, 4-VII-1971, C. D. Ferris (7♂, 1♀ CDF); Jct. of Cassiar Hwy. & Boya Lake Rd., 2200’, 10-VI-1986, C. D. Ferris (12♂, 4♀ CDF); Kelly Lake – Canoe Creek Rd., nr. Jesmond, 3100-4400’, 23-VII-1984, J. & F. Preston (2♀ MGCL); 24-VII-1984, J. & F. Preston (1♂, 1♀ MGCL); [Manning] Park, 5000’, Valley View, 7-V II-1961, H. K immich (1♀ MGCL); Mt. Princeton, 29-30-V-1964, H. Kimmich (1♂ MGCL); Otter Lake C.G., nr. Princeton, 9-VII-1976 (2♂, 1♀ MGCL); Pavilion – Kelly Lake Rd. at Diamond S Ranch, N of Lillooet, 3900’, 24-VII-1984, J. & F. Preston (1♂ MGCL); Pavilion – Kelly Lake Rd., 8.5 mi. N Pavilion, 4300’, 24-VII-1984, J. & F. Preston (2♂, 3♀ MGCL); Pink Mtn., halfway up, on road to lookout, 4-VII-1985, C. D. Ferris (2♂ CDF); Pink Mtn., mi. 147, Alcan Hwy., 9-VII-1978 (2♂ MGCL): Pink Mtn., mi. 147, Hwy. 97, 5000’, VI-VII-1980, N. Tremblay (1♀ MGCL); Princeton, 27-VI-1966 (1♂ MGCL); Princeton, Cardinal Ranch, 27-VI-1966, B. Weber (1♀ MGCL); Stagleap Cyn., Kootenay Dist., 27-VII-1987, D. L. Bauer (1♂ MGCL); Summerland area, Okanagan Valley, 25-VI-1983, J. Reichel (1♂ MGCL); Tompson Mt., Kootenay Dist., 25-VII-1981, D. L. Bauer (2♂ MGCL); 5 mi. S Clinton, 26-VI-1964 (14♂, 5♀ MGCL).
YUKON TERRITORY (317♂, 149♀): Alaska Hwy., bog nr. Johnson’s Crossing, 22-VI-1948 (1♂, 1♀ MGCL); Alaska Hwy., mi. 825, 15-VI-1957, J. & F. Preston (1♂ MGCL); Alaska Hwy., mi. 895-900, nr. Whitehorse, 22-VI-1948 (1♀ MGCL; SN-15-152); Alaska Hwy., mi. 976, nr. Mendenhall, 23-VI-1948 (2♂ MGCL); Campbell Hwy., km. 521, 1800’, 12-VI-1979, J. & F. Preston (1♂ MGCL); Campbell Hwy., km. 533, 1700’, 12-VI-1979, J. & F.Preston (13♂, 10♀ MGCL); Campbell Hwy., km. 563, 1400’, 12-VI-1979, J. & F. Preston (1♂ MGCL); Campbell Hwy., km. 564-568, nr. Carmacks, 62°03’54.24’’N 135°57’00.94”W, 560-650m, 11-VI- 2008, M. Cesanek (15♂, 5♀ MC); Carcross Desert area, Hwy. 2 (Klondike Hwy.), in open woods at desert edge, 2170’, 60°14’14’’N 134°41’41’’W, 29-VI-1985, C. D. Ferris (10♂, 2♀ CDF); Dawson, 13 - VI-1911, ex Barnes coll., “holotype” of “yukonensis” (1♂ MGCL); 14-VI-1911, ex Barnes coll., “allotype” of “yukonensis” (1♀ MGCL); 10 -V I-1981, N. Tremblay (30♂, 8♀ MGCL); Dawson Hwy., mi. 12.6, 16-VI-1962, J. Legge (1♂ MGCL); Dawson-Mayo Loop, mi. 69, W of Whitehorse, 21-VI-1970, D. Eff (2♂ MGCL); Dempster Hwy., mi. 10, 10-VI-1981, J. Johnstone (1♂ MGCL; 1♂ OSUC 618404); 10-VI-1981, N. Tremblay (22♂, 4♀ MGCL; 3♂, 9♀ OSUC 618391- 618399, 618401- 618403); 10 -11-V I-1981, N. Tremblay (2♂ MGCL); 11-VI-1981, N. Tremblay (4♂, 2♀ MGCL; 1♂ OSUC 618379); 19-VI-1981, N. Tremblay (1♀ MGCL); VI-VII-1981, N. Tremblay (3♂ MGCL); 10-VI-1982, N. Tremblay (1♂ OSUC 618428); no date, N. Tremblay (1♂ OSUC 618405); Dempster Hwy., mi. 45- 97, 14-VI-1981, N. Tremblay (1♂ MGCL); Dempster Hwy., mi. 84, 11-VI-1981, N. Tremblay (18♂, 4♀ MGCL); Dempster Hwy., mi. 96, 23-VI-1981, N. Tremblay (2♂ MGCL); Dempster Hwy., mi. 97, 14-VI-1981, N. Tremblay (1♂ MGCL); Dempster Hwy., mi.?, 18-VI-1981, N. Tremblay (1♀ MGCL); 6-VI-1984, N. Tremblay (1♂ MGCL); Haines Jct., 6-VI-1966 (1♂ MGCL); 9-VI-1966 (4♂ MGCL; incl. SN-15-158); 10-VI-1966 (1♂ MGCL); 12-VI-1966 (3♂ MGCL; incl. SN-15-161); 13-VI-1966 (1♀ MGCL); 16 -VI-1966 (1♂, 1♀ MGCL); 17-VI-1966 (1♀ MGCL); 21-VI-1966 (2♂ MGCL); 22- VI-1966 (1♂ MGCL); 24-VI-1966 (1♂, 1♀ MGCL); 25-VI-1966 (4♂, 1♀ MGCL; incl. 3♂ SN-15-160, SN-15-159, SN-15-146); 28-VI-1966 (1♂ MGCL); 29-VI-1966 (9♂, 4♀ MGCL; incl. 1♀ SN-15-148); 1-VII-1966 (1♂, 1♀ SN-15 -169 MGCL); 12-V I-1967 (2♂, 1♀ MGCL); 14-VI-1967 (1♂ MGCL); 18-VI-1967, obtained from J. Ebner (1♂, CDF); 19-VI-1967, J. A. Ebner (1♂ MGCL); 21-VI-1967, J. A. Ebner (1♂ MGCL); 23-VI-1967, J. A. Ebner (1♂ MGCL); 24-VI-1967, J. A. Ebner (1♂ MGCL); 27-VI-1967, obtained from J. A. Ebner (2♀ CDF); 9-VI-1968 (1♂ MGCL); Haines Rd., mi. 87, 30-VI-1966 (1♀ MGCL); Horse Creek, mi. 12.6 Dawson-Mayo Loop, 24-VI-1964, A. H. Legge (2♂, 1♀ MGCL); 24-VI-1964, D. Eff (5♂, 4♀ MGCL); Hwy. 7, Atlin Rd., 2100’, 29-VI-1985, C. D. Ferris (1♂ CDF); Hwy. 11 (Silver Trail), km. 31.5 (SW of Mayo), 27-28-VI-1985, C. D. Ferris (9♂, 3♀ CDF); 4-VI-1987, C. D. Ferris (2♂, CDF); 6-VI-1991, C. D. Ferris (10♂, 3♀ CDF); Jubilee Mtn., 1000-1500m, 5-VII-1977, A. Reif (1♂, 1♀ MGCL); Klondike Hwy., mi. 132, N of Yukon River, 20-VI-1975, D. K. Parshall (1♀ OSUC 618388); Lake Laberge, 30 mi. N Whitehorse, 10-VI-1985, J. & L. Troubridge (4♂ MGCL); 14-VI-1985, T. Kral (4♂ MGCL); 15-VI-1985 (2♂ MGCL); 18- VI-1985, T. Kral (2♂ MGCL); Lake Laberge, Hwy. 2, 46.4 km. N Whitehorse, 11-VI-1981, D. K. Parshall (1♂ OSUC 618439); 1-VII-1981, D. K. Parshall (1♂ OSUC 618442); 1-VII-1982, D. K. Parshall (2♂ OSUC 618390, 618441); 11-VII-1982, D. K. Parshall (1m OSUC 618440); 14 -V I-1983, D. K. Parshall (9♂ OSUC 618327, 618408-618415); 15-VI-1983, D. K. Parshall (1♂ OSUC 618322); 23-VI-1983, D. K. Parshall (3♀ OSUC 618378, 618446-618447); 25-VI-1983, D. K. Parshall (3♀ OSUC 618443-618445); 14-VII- 1983, D. K. Parshall (3♂, 2♀ OSUC 618455-618459); 6-VI-1984, J. P. Ross (2♀ OSUC 618406-618407); 12-VI-1985 (4♂, 1♀ OSUC 618339, 618448-618451); 13-VI-1985, D. K. Parshall (12♂, 5♀ OSUC 618324, 618331-618338, 618430-618437); 14-VI-1985, D. K. Parshall (3♂ MGCL; 13♂, 14♀ OSUC 618323, 618325, 618340- 618354, 618418-618427); 15 -V I-1985, D. K. Parshall (5♂, 5♀ OSUC 618321, 618326, 618328, 618355-618359, 618453-618454); 16 -VI- 1985, D. K. Parshall (4♀ OSUC 618360-618363); 18-VI-1985, D. K. Parshall (6♀ OSUC 618364-618368, 618452); 19-VI-1985, D. K. Parshall (1♂, 4♀ 618369-618373); 20-VI-1985, D. K. Parshall (4♀ 618374-618377); Lake Laberge, Hwy. 2, mi. 29, 3-VI-1985, J. Zeligs (1♂ MGCL); 18-VI-1985, J. Zeligs (1♂ MGCL); Mts. SW of Haines Jct. (5-18 mi.), 3-4000’, 22-VI-1967 (1♂ MGCL); N of Stewart Crossing, Hwy. 2, 22-VI-1983, [D. K. Parshall] (3♂, 5♀ OSUC 618330 [*this specimen with O. tanana barcode, Fig. 10 View Figure 10 c -d], 618380-618386); N of Stewart Crossing, Klondike Hwy., mi. 24, 20 -V I-1975, D. K. Parshall (2♂ OSUC 618329 [*this specimen with O. tanana barcode, Fig. 10 View Figure 10 a -b], 618389); nr. Snafu Lake on Atlin Rd., 2600’, 7-VI-1991, J. & F. Preston (4♂ MGCL); St. Elias Mts., Nickel Ck., 14-VI-1985, B. Grooms (1♂ MGCL); Stewart Crossing, Klondike Loop Rd., 1600’, 13-VI-1979, J. & F. Preston (1♂, 2♀ MGCL); Twin Lakes, Hwy. 2, km. 115, 14-VI-1983, D. K. Parshall (2♀ OSUC 618416-618417); 28-VI-1983, D. K. Parshall (1♂ OSUC 618438); Whitehorse, 10-VII-1919, “paratype” of “yukonensis” (1♀ MGCL); 6 -9 -V I-1923 (1♂ MGCL); 8 -V I-1923 (1♂ MGCL); 9-VI-1923 (2♂ MGCL); 17-VI-1923, J. Kusche (2♂ MGCL); 8-VI-1966 (1♂ MGCL); 9 -V I-1966, H. Ebner (1♂ MGCL); 10 -V I-1966 (1♂ MGCL); 11-VI-1966 (1♀ MGCL); 13-VI-1966 (1♀ MGCL); 2-VI-1982 (1♀ OSUC 618429); 1-VII-1982, B. Grooms (1♂ MGCL); Whitehorse, 2500’, 24-VI-1981, G. Anweiler (1♂ CDF); 8-10-VI-1982, J. P. Ross (3♂, 5♀ CDF); Whitehorse, Baxter coll. (1♂ MGCL); Yukon Hwy. 2 (from Skagway, AK), km. 126, 2550’, 6-VI-1991, J. & F. Preston (5♂ MGCL); 0.8 mi. N of Lewes Lake Rd. on Hwy. 2, 2700’, 6-VI- 1991, J. & F. Preston (6♂, 1♀ MGCL); 1.4 mi. S of Lewes Lake Rd., W of Hwy. 2, 2550’, 6-VI-1991, J. & F. Preston (1♂, 1♀ MGCL); 20 mi. S Burwash Landing, 1-VII-1948 (1♂ MGCL SN-15-150); “Alaska” [old specimen, most likely from Whitehorse area] (1♂ MGCL). Note: Much of the material from the Haines area, including that attributed to J. Ebner, was likely collected by Dr. A. M. Pearson, who collected for Ebner in the Haines area for several years in the 1960’s.
USA: ALASKA: Eagle, 2-VII-1901, S. Hall Young (1♂ MGCL); 27-VI-1903, Reed Heilig, “paratype” of “yukonensis” (1♂ MGCL); Kathul Mtn., N side Yukon River, 6 mi above mouth Kandik R., 29-VI-1975, E. Holsten (3♀ KWP; UAM100379423- UAM100379425); 5-VIII[sic!?]-1975, E. Holsten (1♀ KWP; UAM100379422).
Etymology. This butterfly is named for the Tanana River, which flows through southeastern and central Alaska. Tanana is a Koyukon (Athabascan) word meaning “trail river”, though the term is also applied to an Athabaskan indigenous group (Bright 2004).
Diagnosis. Adults of O. tanana average larger than those of Yukon O. chryxus . The mean forewing length of male O. tanana is 26.7 mm (range 24.6 to 29.4 mm, n = 20), vs. 24.8 mm (range 19.6 to 27.4 mm, n = 20) in Yukon O. chryxus . Females of O. tanana also average larger, with a mean forewing length of 26.9 mm (range 24.9 to 31.3 mm, n = 10), vs. 26.1 mm (range 21.0 to 29.1 mm, n = 10) in Yukon O. chryxus . Adults of O. tanana can usually be identified by the following traits, compared to Yukon O. chryxus : 1) larger size, 2) darker overall upperside coloration, with paler areas dark ochre or reddish, 3) darker ventral forewing coloration, 4) bolder dark ventral hindwing transverse bands, 5) expanded whitish areas on the ventral hindwing, 6) valvae average more robust. While none of these individual characters are strictly diagnostic, when considered together, essentially all specimens can be reliably identified to taxon. In addition, adults of O. tanana are separated from those of Yukon O. chryxus by their unique COI barcode sequences (but see below), which are identical to those found in adjacent populations of O. bore , with the exception of a single base-pair substitution at site 300: G->A (site number corresponds to the sequence given above).
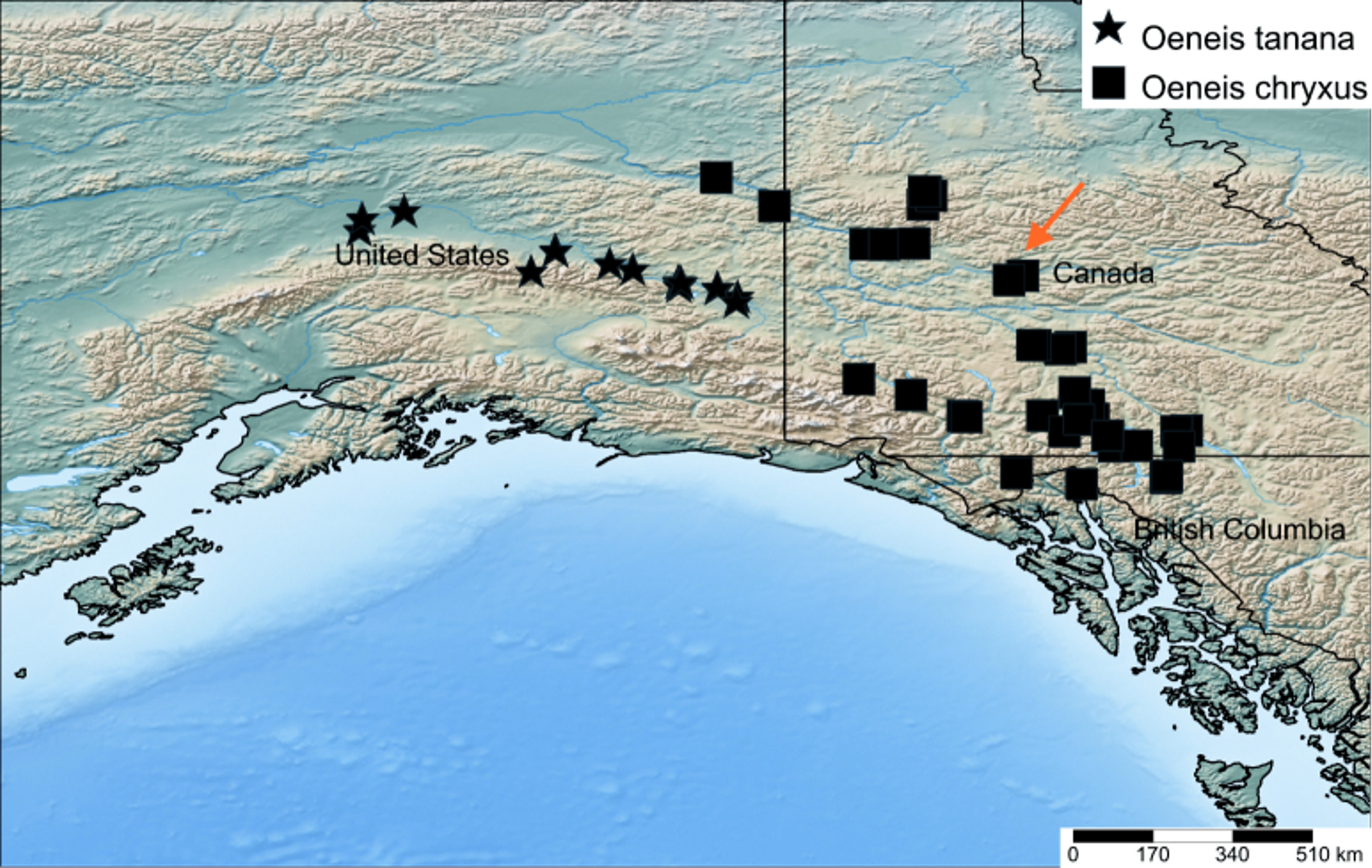
Distribution. All localities where O. tanana is confirmed to occur are within the Tanana River Basin, in southeastern and central Alaska, including the lower north slope of the Alaska Range ( Fig. 7 View Figure 7 ). Available records suggest that O. tanana is widely distributed in appropriate habitats throughout the Tanana River drainage, at least from the Northway area (Northway Airport and Alaska Hwy. mi. 1270), northwest to Nenana, a roughly 400 km. (250 mi.) range centered along the Tanana River. Altitudinal records range from 107 m (351’) at Nenana to 635 m (2083’) along the Richardson Highway (mi. 229) and Delta River, which drains into the Tanana River to the north. A very small part of the Tanana River Basin extends into Yukon Territory (Moran 2007), but it is unknown if O. tanana occurs there. Likewise, it remains unknown how far down the Tanana River Basin O. tanana may occur, or if it occurs along the Yukon River Basin downstream or upstream of its junction with the Tanana River at Tanana . Most of this region has not been surveyed for butterflies. All specimens we have examined of O. tanana were collected in odd-numbered years, with the exception of a single female from Alaska Highway mile 1371, labeled from 1970. No collector’s name is provided on the label, and the possibility of a labeling error cannot be ruled out. The earliest specimen of O. tanana we have seen is from 18 May (1997, 21 mi. SW Fairbanks), and the latest is from 6 July (1995, 5 mi. S of Tok), with most records from the second and third weeks of June.
Habitat. Adults of O. tanana fly in open, dry, grassy areas and clearings in boreal forest. In disturbed areas, they tend to frequent abandoned roads and trails, undeveloped dirt/gravel roads ( Fig. 8 View Figure 8 ), and power line cuts. They are fairly sedentary and in response to a disturbance fly short distances, usually in straight lines, to settle again. The butterflies generally sit on the ground or perch on rocks, or on low vegetation, with wings folded over the back unless basking. While colonies are isolated, numerous individuals are frequently present at occupied sites. Aside from grasses, sedges and various arctic forbs, the principal vegetation at the type locality includes black spruce ( Picea mariana (Mill.) B.S.P.), white spruce ( Picea glauca (Moench) Voss ), quaking aspen ( Populus tremuloides Michx. ), occasional birch ( Betula sp.), and willows ( Salix sp.). Oeneis tanana flies in sympatry with O. jutta (Hübner, [1806]) at Nenana, Alaska Hwy. mi. 1410, and in the vicinity of Tok (including at the type locality), and it flies with both O jutta and O. philipi Troubridge, 1988 at Northway Airport. No information on the early stages or larval foodplants of O. tanana is known to date, although grasses and/or sedges presumably serve as the larval foodplants, as reported for other taxa in the O. chryxus complex (James & Nunnallee 2011).
dI s c u s s I o n
Like O. chryxus , the genitalia of O. tanana possess a tooth-like projection of the dorsal margin of the valva, denticles on the valva in a single series, a strongly sclerotized ventral swelling of the lamella antevaginalis, and a left-skewed vertical plate of the lamella antevaginalis. Lukhtanov & Eitschberger (2001) noted the first three of these characters as diagnostic of the bore group. Based on the presence of these characters, Oeneis tanana is clearly a member of the bore group. Despite the phenotypic differences (adult size and wing color and pattern) between O. tanana and O. chryxus , the genitalia of these two species are very similar. Subtle differences in genitalia of both sexes indicated in Figs. 4-5 View Figure 4 View Figure 5 apparently reflect individual variation. To date, we have not identified any diagnostic characters in the genitalia that serve to unambiguously separate these two taxa, although the valvae of O. tanana average somewhat more robust than those of O. chryxus ( Fig. 6 View Figure 6 ), and are generally slightly larger than those of O. bore . This result is not surprising, considering the lack of consistent genitalic differences reported among other North American members of the bore group. On the other hand, female genitalia of O. tanana and O. chryxus differ from those of O. bore by the position of the vertical projection of the lamella antevaginalis, which is skewed to the left in O. tanana and O. chryxus . Thus, genitalic characters suggest that O. tanana is morphologically closer to O. chryxus than O. bore , although the molecular data discussed below indicate the opposite.
Very little information on O. tanana is available in the literature. We are not aware of any previously published images of adult or immature O. tanana , other than the very recent images of adults of both sexes (as O. chryxus caryi ) by Philip & Ferris (2015). Distributional records for O. chryxus in Alaska provided by Philip (1996, 1998, 2006) and Magoun & Dean (2000) all refer to O. tanana ; we have examined specimens from all but one of these sites. The only molecular study that has focused on the chryxus complex is that by Nice & Shapiro (2001), who studied haplotype variation in 440 base pairs of mitochondrial COII among various western USA populations. Many samples were analyzed from California ( O. c. ivallda and O. c. stanislaus), with others from Idaho, Nevada, Montana, Utah, Colorado, and New Mexico, as well as two specimens from Tok, Alaska (all considered to be O. c. chryxus ). The specimens from Tok (now recognized as O. tanana ) were found to possess a unique haplotype (type ‘E’) not shared with any other populations in the analysis, but no discussion of this population or haplotype was provided.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Satyrinae |
|
Genus |