
Plagioporus, Blend, Charles K., Dronen, Norman O., Gardner, Scott L., Racz, Gabor R. & Armstrong, Howard W., 2012
|
publication ID |
https://doi.org/10.5281/zenodo.208704 |
|
DOI |
https://doi.org/10.5281/zenodo.6168240 |
|
persistent identifier |
https://treatment.plazi.org/id/EB467D70-FFD8-FFC4-C2B2-FF12FED67ED4 |
|
treatment provided by |
Plazi |
|
scientific name |
Plagioporus |
| status |
|
Tellervotrema armstrongi Gibson & Bray, 1982 View in CoL
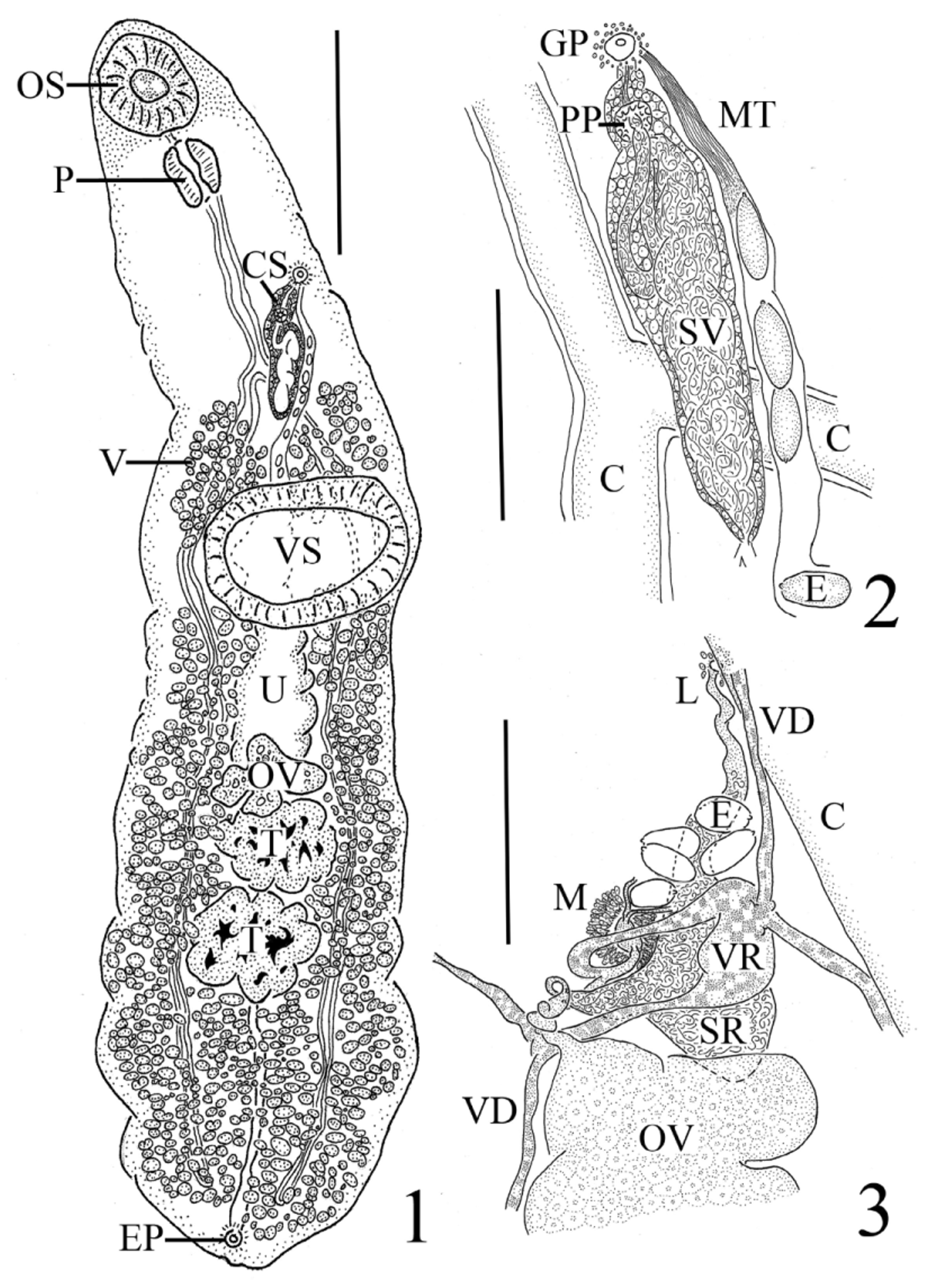
( Figs. 1–3 View FIGURES 1 – 3 )
Synonyms: Plagioporus sp. n. #1 & #2 of Armstrong (1974).
Type-host: Nezumia aequalis (Günther) ; Gadiformes : Macrouridae : Macrourinae ; common Atlantic grenadier.
Other hosts: Nezumia cyrano Marshall & Iwamoto ; Gadiformes : Macrouridae : Macrourinae . Unidentified macrourid; Gadiformes : Macrouridae .
Localities: N. aequalis : Northeastern Gulf of Mexico, 28°02'N, 85°37'W, depth = 585 m, 24/June/1971; 28°15'N, 86°06'W, depth = 589 m, 26/June/1971; 28°22'N, 86°31'W, depth = 713 m; 26/June/1971; 28°31'N, 86°33'W, depth = 534, 27/June/1971; 28°36'N, 86°36'W, depth = 473 m, 27/June/1971; 28°36'N, 86°55'W, depth = 695 m, 28/June/1971; 28°42'N, 86°46'W, depth = 548 m, 28/June/1971; 28°53'N, 86°57'W, depth = 655 m, 29/ June/1971; 28°54'N, 86°55'W, depth = 640 m, 12/February/1970; 28°59'N, 86°49'W, depth = 519 m, 29/June/1971; 29°27'N, 86°57.1'W, depth = 585–732 m, 19/July/1967. N. cyrano: Northcentral and northeastern Gulf of Mexico, 27°31.1'N, 89°48.9'W, depth = 1,064 m, 15/November/1984; 28°06'N, 86°35.3'W, depth = 1,170 m, 18/April/1984. Unidentified macrourid: Northeastern Gulf of Mexico, 29°27'N, 86°57.1'W, depth = 637 m, 19/July/1967.
Site: Intestine; pyloric caecum.
Deposited Specimens: Collector HWA, NMFS & Dr. Thomas Bright, BM(NH) vouchers NHMUK 2012.3.7.1–19 ( 21 adult & 6 immature), USNPC vouchers 105330.00–105335.00 ( 8 adult & 6 immature), HWML vouchers 49746 & 49747 ( 7 adult & 5 immature); Collector LGL, BM(NH) vouchers NHMUK 2012.3.7.20–30 ( 11 adult); Collector Dr. Thomas Bright, BM(NH) voucher NHMUK 2012. 3.7.31 ( 1 adult).
Prevalence: N. aequalis : 19 of 88 host specimens (21.6% infected); N. cyrano : 9 of 14 host specimens (64.3%) infected; unidentified macrourid: 1 of 1 host specimen (100% infected).
Intensity: N. aequalis : 1–11 worms/specimen; N. cyrano : 1–2 worms/specimen; unidentified macrourid: only 1 specimen found.
Mean intensity: N. aequalis : 54 worms in 19 specimens or 2.8 worms/infected fish; N. cyrano : 11 worms in 9 specimens or 1.2 worms/inf. fish; unidentified macrourid: 1 worm in 1 specimen or 1 worm/inf. fish.
Relative density/abundance: N. aequalis : 54 worms in 88 specimens or 0.6 worms/total fish examined; N. cyrano : 11 worms in 14 specimens or 0.8 worms/t.f.ex.; unidentified macrourid: 1 worm in 1 specimen or 1.0 worm/t.f.ex.
Records: 1. Armstrong (1974); 2. Gibson & Bray (1982); 3. Bray (1995); 4. Blend (1996); 5. Klimpel et al. (2001); 6. Bray & Kuchta (2006); 7. Klimpel et al. (2009); 8. Overstreet et al. (2009); 9. Present study.
Descriptions: 1, 2, 4, 9.
Redescription. [Based on 48 specimens. Measurements and proportions are given in Table 2.] Body elongateoval, widest postequatorially; lateral margins occasionally foliate or crinkled, especially in posterior region. Forebody tapered anteriorly. Hindbody either slightly truncate or plump and rounded posteriorly. Tegument smooth. Pre-oral lobe absent. Oral sucker spherical to subspherical, subterminal. Ventral sucker sessile, transversely elongate or oval, wider than long, larger than oral sucker, preequatorial. Prepharynx quite short. Pharynx muscular, oval, longer than wide. Oesophagus distinct, with tegumental lining, straight in most individuals (92%), slightly convoluted rarely in others (8%). Intestinal bifurcation located 3/4 to 4/5 distance between anterior extremity and ventral sucker. Caeca narrow, unbranched, with thickened walls, ending blindly near posterior extremity.
Testes 2, lobed or irregularly indented or smooth, subspherical to transversely elongate or oval, tandem, median, intercaecal, postequatorial in posterior third of body, contiguous in most individuals (58%) but can be separated by short distance (42%). Post-testicular region fairly sizeable, occupying posterior third of body. Cirrus-sac thin-walled, clavate, straight, entirely preacetabular (can extend to midacetabular level in contracted specimens), extending posterodextrally from point midway between left lateral body edge and midline to terminate at or just posterior to level of intestinal bifurcation near midline. Seminal vesicle bipartite; posterior portion saccular and occupying most of cirrus-sac; anterior portion tubular, looping over posterior portion before extending anteriorly in to narrow, distal portion of cirrus-sac to terminate in small genital atrium. Prostate gland-cells well developed, numerous in cirrus-sac. Pars prostatica, narrow ejaculatory duct, and cirrus present. Genital pore submedian, sinistral, midway between body margin and midline but can be nearer to lateral margin in some individuals, prebifurcal in region between mid-oesophagus and intestinal bifurcation, preacetabular and midway between oral and ventral suckers; dark-stained cells surrounding pore.
Ovary deeply 3- to 4-lobed and clover-leaf-shaped, median, intercaecal, immediately pretesticular, contiguous with anterior testis, postequatorial and located near junction of middle and posterior thirds of body. Oviduct arises anteriorly from anterodextral lobe of ovary. Oötype located on median line or immediately anterosinistral and dorsal to ovary. Seminal receptacle canalicular, ovoid to teardrop-shaped, dorsosinistral and either completely anterior to or overlapping anterior margin of ovary. Laurer's canal wide and winding, passing anteriorly from seminal receptacle to overlap posterosinistral loops of uterus, terminating directly dorsal to or just medial of left cecum on dorsal surface; dark-stained cells around canal opening. Vitellaria follicular, circumcaecal, extending posterolaterally from level at or just posterior to intestinal bifurcation to posterior extremity; vitelline field interrupted around level of ventral sucker producing two paired, non-confluent, isolated patches of vitelline follicles extending from bifurcal level to anterior margin of ventral sucker (paired anterior vitelline patches can be confluent and extend 3/4 distance over ventral sucker in contracted specimens); remainder of vitelline follicles commencing at level of posterior margin of ventral sucker, running posterolaterally to testes and ovary, not confluent in immediate preovarian space, between ovary and anterior testis or in intertesticular area, confluent in posttesticular region. Vitelline reservoir medial to sinistral, dorsally overlapping anterior margin of ovary or immediately preovarian, often ventrally overlapping seminal receptacle. Uterus tightly coiled, winding, intercaecal, wide loops running anteriorly from ovary up through middle of ventral sucker where loops narrow; distal loop turning anterosinistrally to run up along left side of cirrus-sac before terminating at genital pore. Metraterm straight, thin-walled, weakly developed, narrower than remainder of uterus, 1/5 to 1/3 length of cirrus-sac and running anterior along left side of it. Eggs smooth, oval, partially to severely collapsed in many specimens, yellow, non-filamented, operculate, with conspicuous knob or boss on one pole.
Excretory vesicle tubular, I-shaped, terminating at ovary or in region between ovary and anterior testis. Excretory pore dorsal, subterminal, opening via a small sphincter; elongated, dark-stained cells surrounding pore.
Remarks: With the presence of a well-developed cirrus-sac enclosing a seminal vesicle and a canalicular seminal receptacle, our specimens belong within the opecoelid subfamily Plagioporinae . Based on the key by Cribb (2005), these same specimens can be identified as belonging to the genus Tellervotrema , distinguished from other plagioporines by the presence of the following combination of diagnostic characteristics: non-filamented eggs that are> 40 µm long (ours ranged from 47.5–66 µm long); blind caeca; an elliptical, non-pedunculate ventral sucker that lacks lamellar lips, “fleshy folds” or an accessory attachment organ; vitelline follicles that extend to the posterior end of the body, well into the forebody, and are distinctly interrupted at the level of the ventral sucker; tandem and paired testes; an oral sucker that is not funnel-shaped; an excretory vesicle that is not diverticulate and does not extend to the pharynx but terminates inside the hindbody at about the level of the ovary; a clearly submedian genital pore; a pre-testicular uterus; and our specimens were found in macrourid hosts.
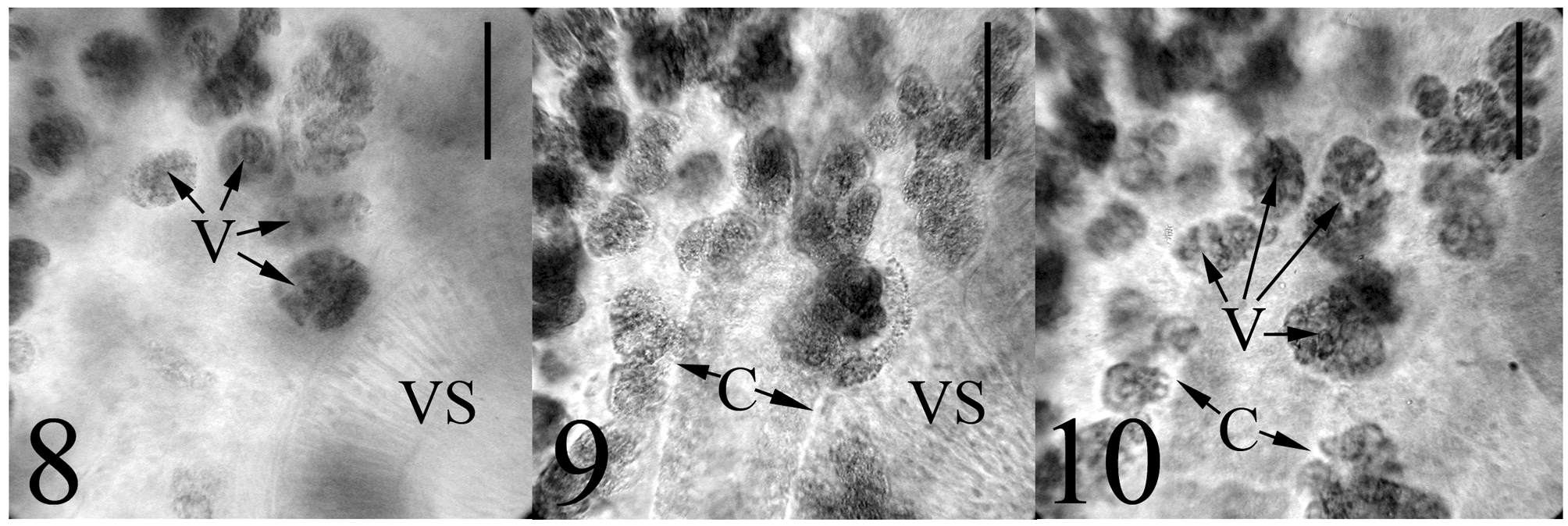
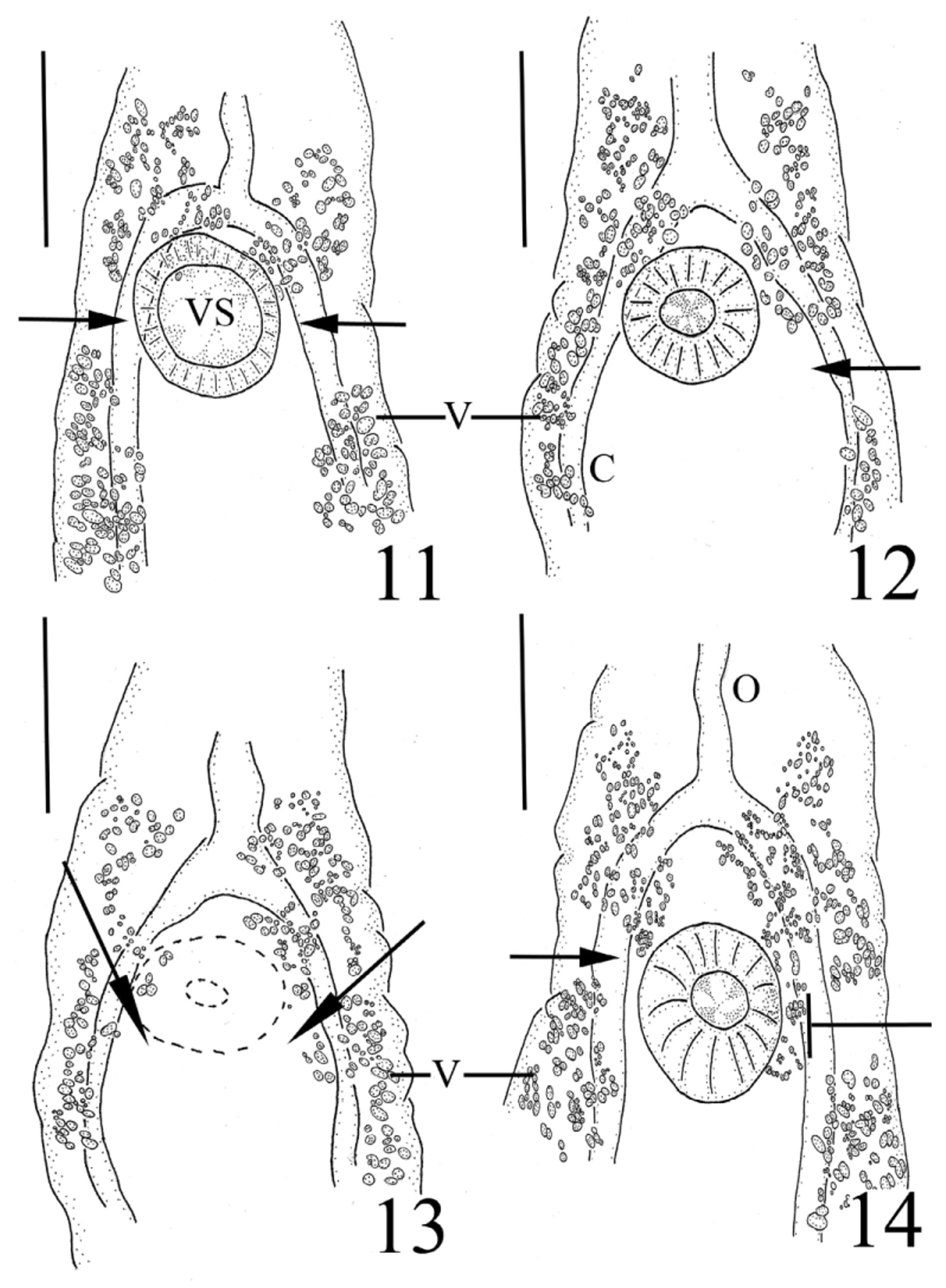
Our specimens compare favourably to T. armstrongi as described by Gibson & Bray (1982) in almost every aspect including overall appearance, the presence of a symmetrical pair of isolated groups of follicles in the posterior forebody proximate to the ventral sucker, a sinistral genital pore, a conspicuous trilobed ovary (4-lobes also observed, see Figs. 18–19 View FIGURES 15 – 19. 15 ), tandem and paired testes that may be contiguous or separated by a short gap, a unique dorsally subterminal excretory pore opening via a sphincter surrounded by gland cells, and our specimens were obtained from the intestine of two nominal species of macrourid hosts (the type host species, N. aequalis , and N. cyrano ) as well as from an unidentified macrourid species. We noted a few differences between the description of the type specimens of T. armstrongi and our specimens. Tellervotrema armstrongi was described as possessing inpart the following characteristics: a distinct prepharynx which could reach up to half the length of the pharynx; irregularly lobed testes; and vitelline follicles that do not occur dorsally to the caeca ( Gibson & Bray 1982). Observations of our specimens revealed more variability in the first two characteristics mentioned immediately abovewhile some individuals possessed a distinct, short prepharynx ( Fig. 1 View FIGURES 1 – 3 ), we were not able to observe this feature in others (0-88 µm long), and we noticed a breadth of testes textures, ranging from smooth to irregularly indented to lobed (see Figs. 20 & 21 View FIGURES 20 – 23 ). Most significant was the presence of circumcaecal vitelline follicles in our material. While there is a tendency for the more posterior vitelline follicles to be restricted to only the ventral side of the caeca, we noticed at times circumcaecal vitelline follicles in this location. More numerous were the circumcaecal follicles making up the two isolated anterior vitelline groups (see Figs. 8–10 View FIGURES 8 – 10 ). Gibson &Bray (1982) noted that species of this genus have vitelline follicles that do not occur dorsally to the caeca and they illustrated this in Figure 12 View FIGURES 11 – 14 B–C. However, these transverse sections were taken from the levels of the anterior testis and the middle of the posttesticular region, respectively—levels posterior to the two anterior isolated vitelline groups characteristic of Tellervotrema . Had sectioned material been illustrated in this anterior region of the worm, the circumcaecal nature of the vitelline follicles might have been more evident.
This report includes the original specimens described and illustrated as “ Plagioporus sp. n. #1 & #2” by one of us (HWA) (see Armstrong 1974, p. 82–89). We have confirmed the speculation noted earlier by Gibson & Bray (1982, p. 551) that these two species of Plagioporus are indeed conspecific and represent T. armstrongi . “ Plagioporus sp. n. #2” is a contracted form of “ Plagioporus sp. n. #1” (= T. armstrongi ). This is evident in Table 2 by the smaller size ranges of several features for these three specimens of T. armstrongi . The host species and geographic locality for “ Plagioporus sp. n. #1 & #2” were the same—the type host, N. aequalis , and the northeastern Gulf of Mexico. Finally, the three specimens of “ Plagioporus sp. n. #2” were found in co-infection with one specimen of “ Plagioporus sp. n. #1” ( Armstrong 1974, p. 89).
This account represents a new host record in that this is the first original report of T. armstrongi from the macrourid N. cyrano . This digenean species has already been documented in the northeastern Gulf of Mexico ( Gibson & Bray 1982).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Plagioporinae |