
Neoperla coreensis Ra, Kim, Kang & Ham, 1994
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3918.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:392562D1-8772-4F8D-885C-57CAA5B670C1 |
|
DOI |
https://doi.org/10.5281/zenodo.6095852 |
|
persistent identifier |
https://treatment.plazi.org/id/9A3687BC-5D24-5600-FF76-8A61FD72B708 |
|
treatment provided by |
Plazi |
|
scientific name |
Neoperla coreensis Ra, Kim, Kang & Ham, 1994 |
| status |
|
Neoperla coreensis Ra, Kim, Kang & Ham, 1994 View in CoL
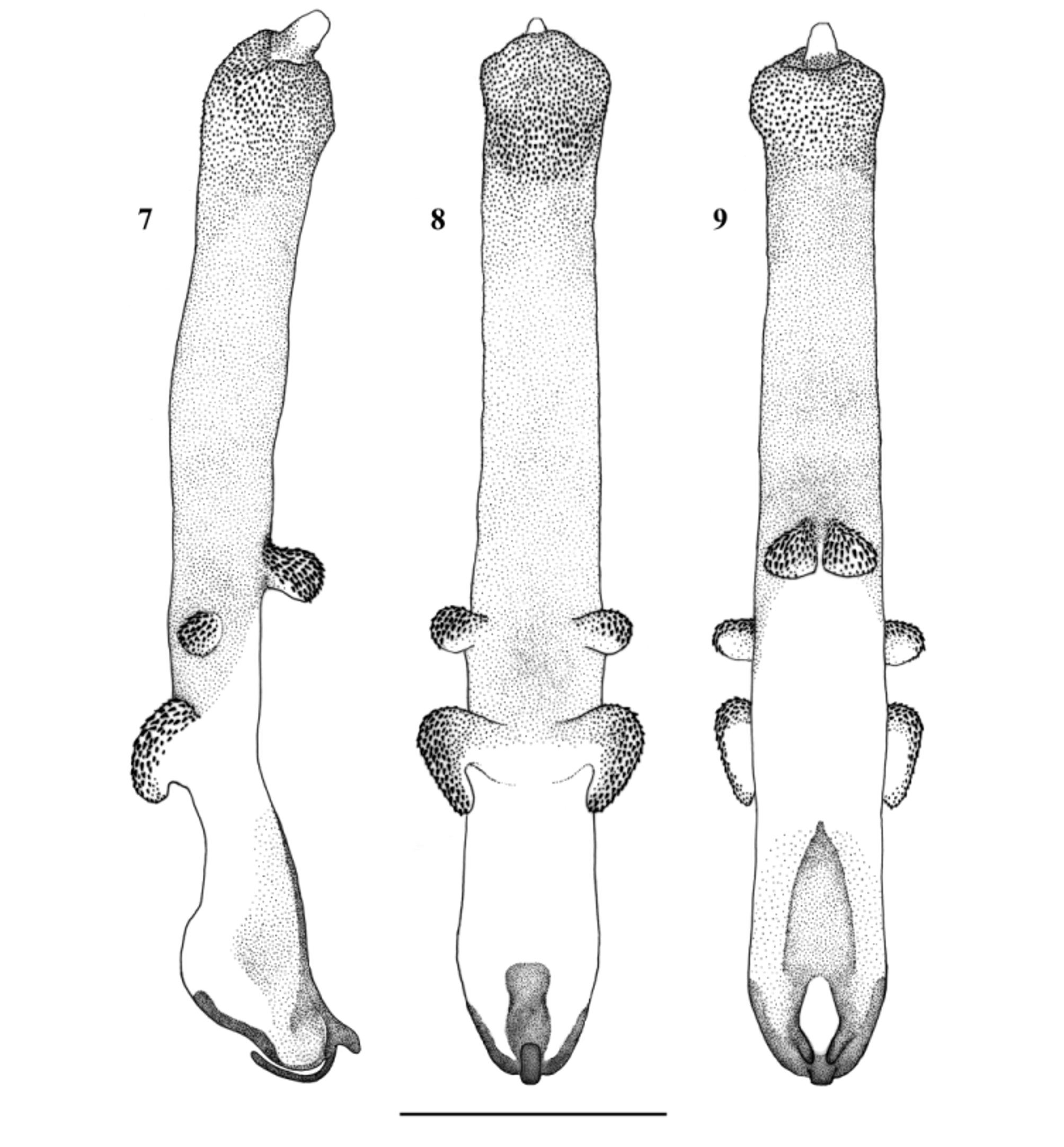
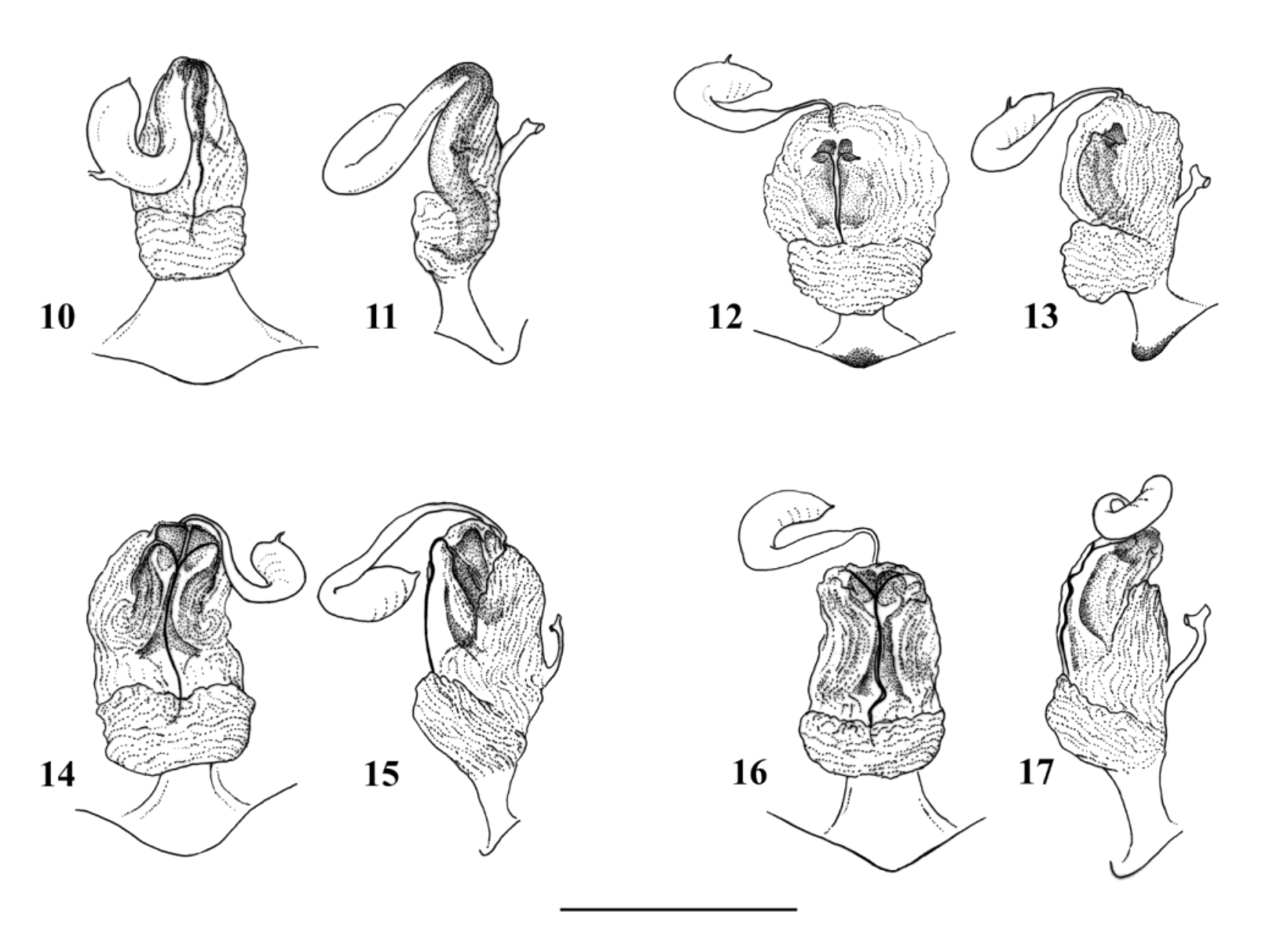
( Figs. 7–9 View FIGURES 7 – 9 , 12–13 View FIGURES 10 – 17 , 24–28 View FIGURES 24 – 33 , 41 View FIGURES 41 – 42 )
Type locality: Korea, Kwangju, Mt. Mudung, Sanjang Valley ( South Korea, Gwangju Metropolitan city, Mt. Mudeung, N35°10’ E126°55’).
Neoperla coreensis Ra, Kim, Kang & Ham, 1994 View in CoL — Ra, Kim, Kang & Ham 1994: 9. (original description of male, female and larva); DeWalt et al. 2014 (catalog).
Literature data. Ra et al. (1994): South Korea, Kwangju, Mt. Mudung, Sanjang Valley (Gwangju Metropolitan city, Mt. Mudeung, N35°10’ E126°55’), 06.VII.1993, leg. Jong Sun Kim: holotype ♂, 24♂ 10♀ and 1 nymph paratypes; same locality, 26.VI.1993, leg. Jong Sun Kim, Soon Ah Ham: 2♂ 2♀ and 1 nymph paratypes; same locality, 17.VI.1993, leg. Jong Sun Kim: 1♂ 1♀ paratypes.
Material examined. NORTH KOREA: North Pyongan Province, Hyangsan-gun, Myohyang Mts., at light on balcony and garden of Hotel Myohyang-san (locality No. 793), 150 m a.s.l., N40°00’ E126°15’, 14.VII.1982, leg. László Forró, László Ronkay: 1♀ ( HNHM; eggs prepared for SEM, terminalia cleared in KOH and stored in the same vial); North Pyongan Province, Hyangsan-gun, Myohyang Mts, Hotel Myohyang-san, 150 m a.s.l., N40°00’ E126°15’, 12.VIII.1989, leg. Lajos Berczi: 1♂ ( HNHM; terminalia and aedeagus cleared in KOH and stored in the same vial).
Complementary description. Aedeagus ( Figs. 7–9 View FIGURES 7 – 9 ): The aedeagal tube plump, with a triangular dorsobasal sclerite, a ventrobasal hump and two large, elongated ventroapical lobes. Dorsal and dorsolateral surface covered with fine spinules, other surfaces of the tube without spines. The ventroapical lobes backcurved, not touching each other, and bear large spines on their ventral surface, dorsally without spines. The everted aedeagal sac straight, 2× as long as the tube. Basal ¼ of the sac with a pair of small, hemispherical lobes lateroventrally and a more apically positioned, dorsal pair of larger and more elongated lobes. Both pairs of lobes covered with large spines, but basal surface without spines. Sac covered with fine spinules to the apex, the slightly bulging apical ¼ armed with stronger spines. The separated, slightly downcurved apex of the sac without spines.
Female terminalia ( Figs 12–13 View FIGURES 10 – 17 ): Sterna and terga simple, posterior edge of S8 forms a small, triangular subgenital plate distinctly darkened. Vagina large, nearly as long as wide and with many membranous folds. Inner sclerite large, ending in two small lobes dorsally to anterior folds, basal portion plate-like. The inner pair of lateral folds bear dark, well sclerotized crest short and scarcely reaching apical ones of the posterior folds. Spermatheca small, its origin covered by protruding lateral folds in dorsal view.
Egg ( Figs 24–28 View FIGURES 24 – 33 ): Chorion dark brown, 0.38–0.42 mm long and 0.16–0.22 mm wide (N=10). Barrel-shaped, cross section round. Hatching line inconspicuous. Micropyles placed in a transverse row closer to the opercular end, each located between striae. Collar sessile without distinct flange, surrounded by three to four rows of regular FCIs that are with scarce inner punctation; anchor not studied. Chorion covered with striae grouped only before collar and lid end; outer striae slightly raised but not thickened. Striae set closely without sulci between them. Lid bears five to six distinct rows of FCIs at junction of striae, only the apical one or two rows are with less distinct FCIs; all are with fine inner punctations.
Remarks. This is a member of the montivaga group, but no closely related species is evident. The aedeagal tube of N. magisterchoui Du, 2000 is similar to the uneverted aedeagus of N. coreensis , but the aedeagal sac is different. Possibly, certain Philippine species with very complex aedeagal lobes ( Sivec 1984) may be related to this distinct taxon. Development of the vagina is typical for the lushana subgroup, can be distinguished from those of the known females by the short but distinct crest of inner pair of lateral folds, and wide portion of the lateral folds. The egg is typical Neoperla type, scarcely distinguishable from several other species, e.g. the sympatric N. goguryeo . The female was associated with the male on the basis of similar coloration, and darkened subgenital plate depicted in the original description ( Ra et al. 1994).
Distribution and ecology. The species was described from Mt. Mudung that is part of the Sobaek Ranges of the southern Korean Peninsula. This species has not been reported since the original description. Our specimens were collected at lights in the lower region of the Myohyang Mountains, together with N. goguryeo . Its occurrence in the northern part of the Peninsula assumes a wide distribution of the species, at least in Korea ( Fig. 41 View FIGURES 41 – 42 ). It is probably a species of large streams and small rivers.


| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Neoperla coreensis Ra, Kim, Kang & Ham, 1994
| Murányi, Dávid, Li, Weihai, Jeon, Mi Jeong, Hwang, Jeong Mi & Seo, Hong Yul 2015 |
Neoperla coreensis
| Ra 1994: 9 |