
Neomaenas monachus monachus ( Blanchard, 1852 ) (Satyrus)
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4125.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:118F4865-D89E-45EA-A210-8D61946CC37F |
|
DOI |
https://doi.org/10.5281/zenodo.6070042 |
|
persistent identifier |
https://treatment.plazi.org/id/03F187D7-FFAE-845A-FF11-FCF4FBECB804 |
|
treatment provided by |
Plazi |
|
scientific name |
Neomaenas monachus monachus ( Blanchard, 1852 ) (Satyrus) |
| status |
|
Neomaenas monachus monachus ( Blanchard, 1852) (Satyrus) View in CoL comb. rev.
( Figs. 7 View FIGURE 7 C; 15M–O; 26)
Lectotype: (male) MNHN, Paris, Genitalia P. Viette, Prep. no 4144 (Photo examined)
Type location: Chile
= Epinephele valdiviae C. Felder & R. Felder, 1867
Lectotype: (male) BMNH #809780 (specimen examined)
Paralectotype: (male) BMNH #809781 (specimen examined)
Type location: Valdivia, Los Rios province, Chile
= Pedaliodes lugubris Butler, 1870 View in CoL
Type location: Chile, probably Valdivia, Los Rios Province, Chile
Holotype: (male) BMNH #809743 (specimen examined)
= Satyrus luctuosus Reed, 1877
Holotype by indication ( ICZN Art. 12.2.7): Plate II, Fig. 5 View FIGURE 5 of Reed (1877)
= Satyrus monarchus Reed, 1877 (misspelling)
= Maniola monachus sacerdos Bryk, 1944
Holotype: (male) NRM, Stockholm (photo examined)
Allotype: (female) NRM, Stockholm (not examined)
Paratypes (11 males, 4 females) NRM, Stockholm (not examined)
Type location: Peninsula Llau Llau, north of Punto Nuevo, Lago Nahuel Huapí, Neuquen Province, Argentina, Oct. 1933 - March 1934, leg. Anna Hildegard Ljungner.
Subspecies:
Neomaenas monachus limonias ( Philippi, 1859) (Satyrus) n. comb. (see below)
Other combinations:
Elina monachus — Heimlich (1972)
Quilaphoetosus monachus View in CoL —Herrera (1966); Lamas & Viloria (2004, p. 217)
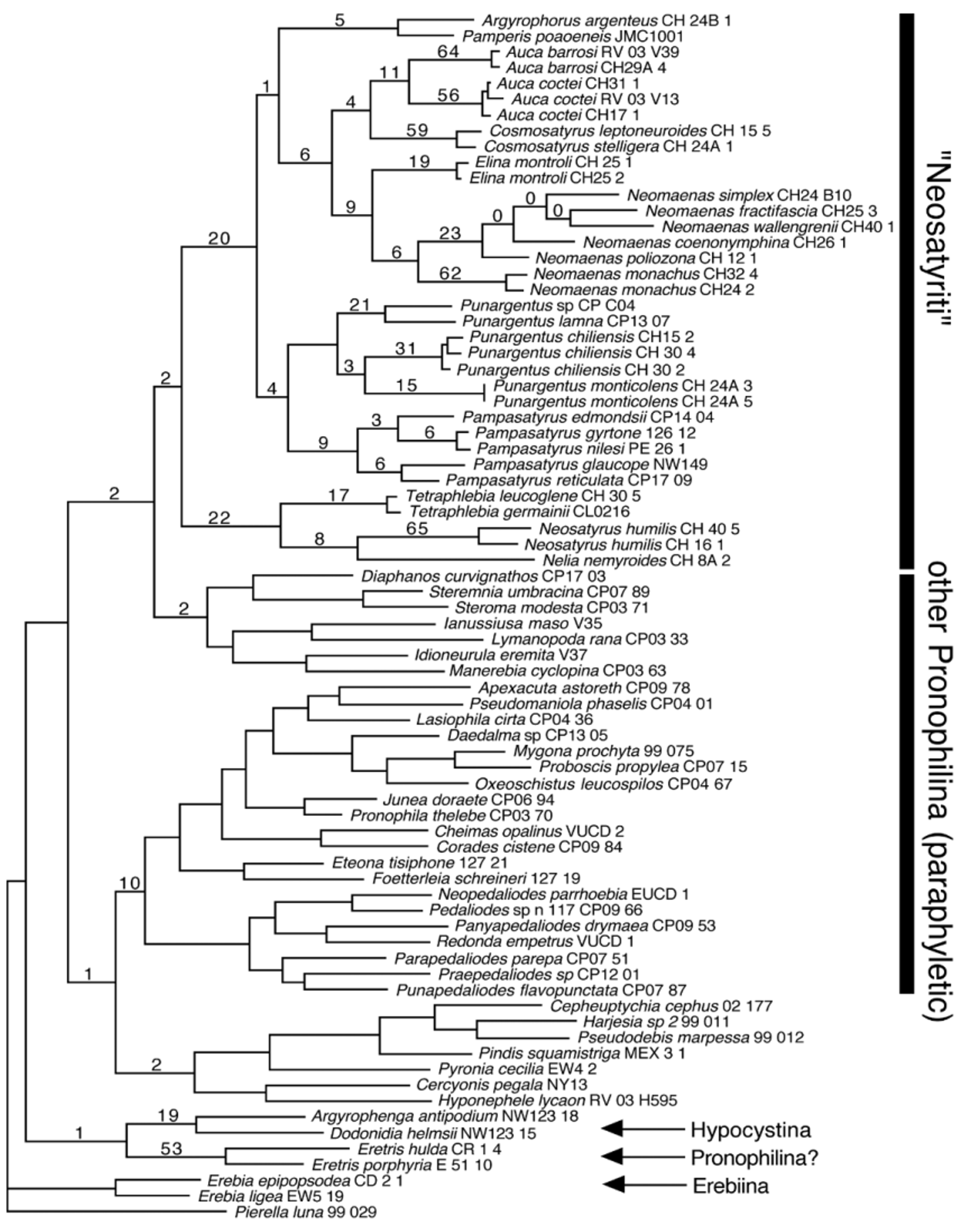
Distribution. Found in Chile from Santiago, Metropolitan Province to northern Los Lagos Province, and in Argentina in Southern Neuquén Province from mid-December to early March at nearly sea level to 1850m ( Fig. 26). Adults can be found flying around the tops of tall stands of bamboo.
Diagnosis. Most similar to Auca coctei , but tending to be larger and darker in color. Males with heavy androconia in the cells between M3 and the inner margin. Ocellus between M1-M2 on the ventral side of the forewing is usually unipupillate. Postmedian band similar to A. coctei , but with both subterminal and median borders deckle-edged. Small, round ocelli are present in each cell between Rs and 1A+2A, the pairs closest to the costa and closest to the inner margin may be oval and black, ringed in cream to daffodil yellow while the center pair are usually white to daffodil yellow. Hindwing ocelli are sometimes obscured or absent. Eyes are hairy and foreleg tarsi of both sexes are unsegmented and club-like. Male genitalia appear more elongated in the uncus and gnathos and wider in the valvae than in A. coctei , but are otherwise similar.
Redescription. Head: Antennae 8–10mm, covered in dark chocolate scales with a longitudinal daffodil yellow stripe, and terminating in a spatulate club. Eyes hairy and oval, length approximately 1.3 times width. Palps with a longitudinal black stripe bordered in white at the median, dark chocolate dorsally and dark chocolate and white ventrally, the terminal segment entirely dark chocolate. Females the same, but lighter in color. Terminal palp segment cylindrical and a little less than one-third the length of the second segment.
Thorax dark amber, males with iridescent black scales and tawny and black piliform scales. Females with white and iridescent black scales and tawny piliform scales. Male abdomens coffee-colored on the dorsal side and coffee-colored with sparse white and tawny scales on the ventral side. Females coffee dorsally and cream to taupe ventrally. Forelegs of both sexes with the tarsus club-like, unsegmented, and without spines. Midlegs and hindlegs with four rows of black spines on the tibia and tarsus.
Forewing ( Fig. 7 View FIGURE 7 C): Wingspan 24–32mm. Males with androconia in chevron to rectangular patches in each cell between M3 and the inner margin. Termen nearly straight to slightly concave and barely scalloped. The distal end of the discal cell is sinuous with the costal half more deeply curved than the cubital half. Dorsal side dark chocolate brown to dark coffee brown with fringe scales in dark chocolate and white to cream, giving the appearance of crenulation. A reddish patch may be visible from the discal cell to the median and a round, black apical ocellus is usually visible between M1-M2. Ventral side chocolate to dark chocolate with a ripple pattern in dark chocolate to dark coffee striations extending from the costa to the radials and from the middle of the postmedian band to the termen. A rust orange to rust red patch extends from the discal cell to the postmedian band, covering the median and cubital veins, extending almost to the anal vein. Postmedian band is lightly bordered in dark chocolate to dark coffee, the median and subterminal sides deckle-edged. Rust orange to rust red patch lightens slightly inside the postmedian band. Apical ocellus between M1-M2 is round, black, unipupillate, and ringed in rust orange to daffodil yellow. This ocellus occasionally extends from M1-M3 and may be bipupillate. Females are lighter in color than males.
Hindwing ( Fig. 7 View FIGURE 7 C): Wing trapezoidal, termen slightly convex and scalloped, inner margin excavated between the anal vein and 1A+2A. Dorsal side and fringe scales similar in color to the forewing, the postmedian band sometimes appearing between M2 and CuA 2 in triangular patches. Ventral side chocolate to dark chocolate brown with a ripple pattern in dark chocolate to dark coffee over the entire wing. Postmedian band lighter than the rest of the wing, with both median and subterminal borders deckle-edged, and wider over the radials than from Rs to the inner margin. Ocelli between Rs-M1, M1-M2, CuA1-CuA2, and CuA2-1A+2A are small and either oval and black, ringed in cream to daffodil yellow or round and white to daffodil yellow. Ocelli between M2-M3 and M3-CuA1 are small, round, and white to daffodil yellow.
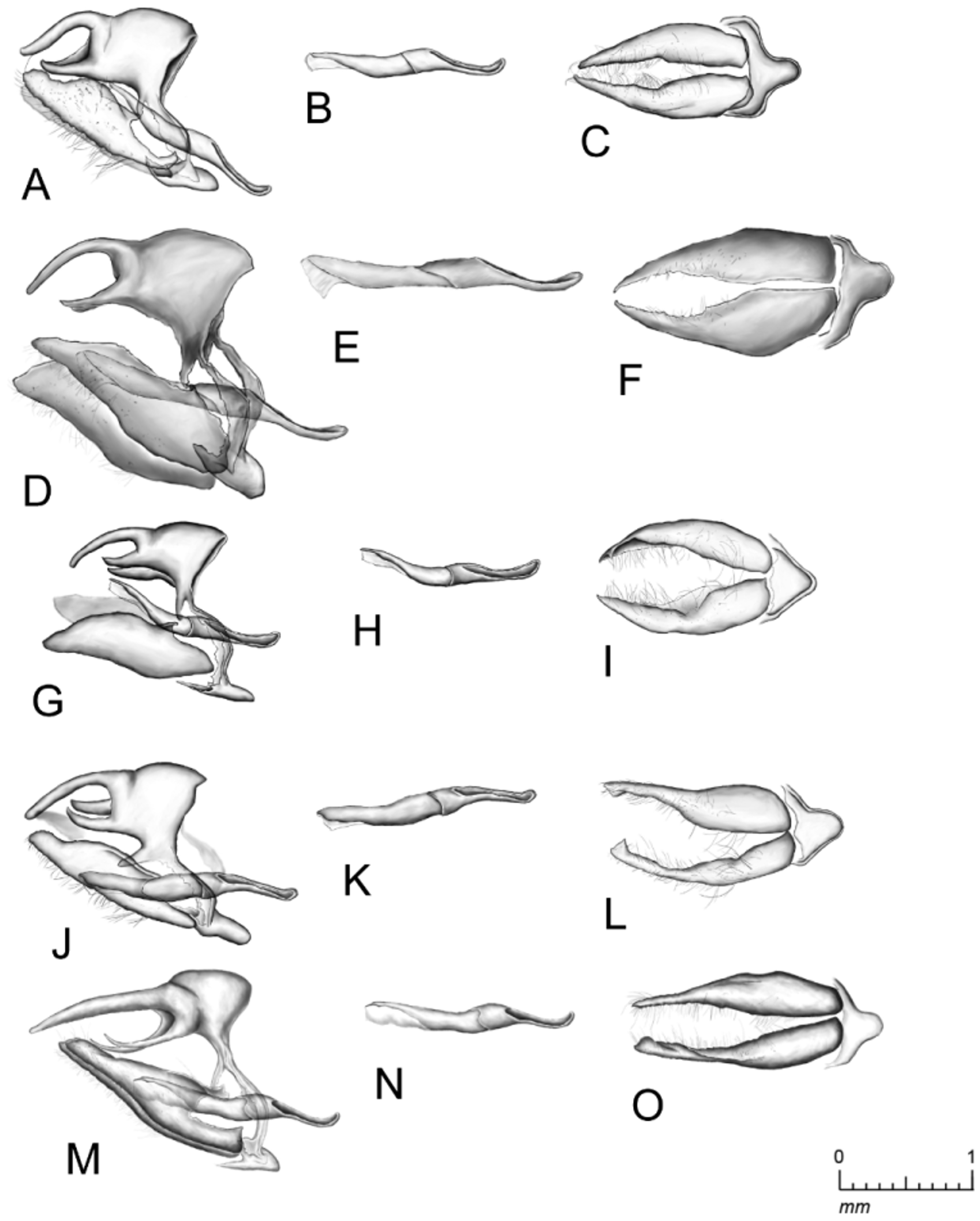
Male genitalia ( Fig. 15 View FIGURE 15 M–O): Uncus widest at the base, narrowing gradually to a blunt finger-like terminus, and approximately 1.25 times as long as the tegumen. Gnathos acute and a little more than half the length of the uncus. Pedunculus long and U-shaped. Saccus U-shaped and approximately four-tenths the length of the gnathos. Valvae widest at the proximal one-quarter, gradually narrowing toward the distal end, the terminus deltoid or somewhat falcate. Aedeagus hourglass-shaped at the distal three-fifths, narrowing slightly to a U-shaped proximal terminus.
Remarks. Originally placed in Satyrus , this species was placed under the generic name Quilaphoetosus by Herrera (1966), based primarily on subtle genitalic differences and variation in the position of the forewing radial veins, a feature that varies widely amongst individuals of the same species collected at the same locality. With the exception of the autapomorphic androconial patch in the males and densely hairy eyes, it is very similar to other members of Neomaenas , where Hayward (1958), D’Abrera (1988) and Peña & Ugarte (1997) had placed it. Neomaenas monachus is the sister taxon to other Neomaenas species sampled in our phylogenetic analysis ( Fig. 1 View FIGURE 1 ), and although these autapomorphies could be used as evidence to argue for the retention of Quilaphoetosus while maintaining an otherwise monophyletic Neomaenas , that genus would be monotypic, and we feel that taxonomy is better served by generic names that imply groups.
Elwes (1903) claimed that it is “very abundant in the heavy virgin forest south of Temuco” and “amongst the dense bamboo undergrowth in the Upper Renaico Valley.” N. monachus monachus may also be found in Chubut, Neuquén, and Rio Negro Provinces, Argentina ( Butler 1881, Elwes 1903). This species tends to fly over the tops of Chusquea bamboos near forested areas, at a height of about 2.5–3m above the ground.
Specimens examined. Chile, Bío-Bío Province, (OSU) 0 0 0 0 94359, 0 0 0 0 94363, 0 0 0 0 94373, (MTSU) CH 24- 1- CH 24-5, (UJ) 1 female; Chile, Araucanía Province, (MTSU) CL0810-CL0814, CL0721-CL0724(UJ) 9 males, 3 females; Chile, Los Lagos Province, (OSU) 0 0 0 0 94377, 0 0 0 0 94372, 0 0 0 0 94362, 0 0 0 0 94375 (MTSU) CH 10-1- CH 10-3, CH 10-8, CH 10B-10, CH 10B-11, CH 10B-2, CH 10B-6, CH 10C-3, CH 7-1


| NRM |
Swedish Museum of Natural History - Zoological Collections |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Satyrinae |
|
Genus |
Neomaenas monachus monachus ( Blanchard, 1852 ) (Satyrus)
| Matz, Jess & Brower, Andrew V. Z. 2016 |
Maniola monachus sacerdos
| Bryk 1944 |
Satyrus luctuosus
| Reed 1877 |
Satyrus monarchus
| Reed 1877 |
Pedaliodes lugubris
| Butler 1870 |
Epinephele valdiviae
| C. Felder & R. Felder 1867 |
Neomaenas monachus limonias (
| Philippi 1859 |