
Myrmekioderma intrastrongyla, Sandes, Joana & Pinheiro, Ulisses, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3702.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:06CCDF7C-86D4-4711-9E42-C9EA4AB02951 |
|
DOI |
https://doi.org/10.5281/zenodo.5674189 |
|
persistent identifier |
https://treatment.plazi.org/id/1E442F3C-FFA2-FFC5-FF18-F8F5FC2DFD3D |
|
treatment provided by |
Plazi |
|
scientific name |
Myrmekioderma intrastrongyla |
| status |
sp. nov. |
Myrmekioderma intrastrongyla sp. nov.
( Figs. 2–3 View FIGURE 2 View FIGURE 3 )
Type Specimens: Holotype – MNRJ 16784. off Aracaju (11º03’14.71’’S 36º54’52.36’’ W), Sergipe State, Brazil, depth 30 m, leg. Petrobras (06/2002). Paratype: UFPEPOR1528 and UFSPOR184, off Aracaju (11º03’14.71’’S 36º54’52.36’’ W), Sergipe State, Brazil, depth 30 m, leg. Petrobras (06/2002); UFPEPOR1529, UFSPOR137 and UFSPOR160, off Pirambu (10º45’36’’S 36º36’08’’W), Sergipe State, Brazil, depth 20 m, col. Cosme Assis and Damião Assis (06/2003). UFPEPOR1530 and UFSPOR188, off Piaçabuçu (10º24’30.95’’ S 36º03’6.55’’ W), Alagoas State, Brazil, depth 30 m, leg. Petrobras (12/ 2002 and 06 /2003, respectively).
Diagnosis. Myrmekioderma with choanosomal strongyles, only one category of smaller acanthoxea and two size categories of trichodragmata.
External morphology ( Fig. 2 View FIGURE 2 ). Massive form, the holotype with 8 x 10 x 2 cm (length x width x height) and paratypes 4.5–8.5 x 2.5–4.5 x 1 –3.5 cm (length x width x height) ( Fig. 2 View FIGURE 2 A–C). The specimens are fragmented. The oscules are circular and can only be observed in the inner part of the fragments. The surface characteristics vary being composed of polygonal protuberances (honeycomb) separated by shallow grooves ( Fig. 2 View FIGURE 2 C), with some regions warty, or tubercular projections ( Fig. 2 View FIGURE 2 A–B). Consistency is slightly compressible. Color pinkish in spirit.
Skeleton ( Fig. 3 View FIGURE 3 A–B). Ectosomal skeleton a detachable peel, formed by acanthoxeas in palisade, crossing the surface ( Fig. 3 View FIGURE 3 A). Choanosome cavernous, composed by acanthoxeas, trichodragmata and strongyles. In its most superficial part, some ascends multispicular tracts through the ectosome, in contrast to a confused central region, which the spicules are disorganized. Besides of that, it also has a subectosomal region, formed by spicules disorganized in high concentration ( Fig. 3 View FIGURE 3 A–B).
Spicules ( Fig. 3 View FIGURE 3 C–M; Tab. 1 View TABLE 1 ). Smooth strongyles located only in the choanosome, larger than the oxeas, measures 300–550.2–792/6–11.1–20 µm, rarely with stylote modification ( Fig. 3 View FIGURE 3 C–D). Acanthoxeas slightly curved in the center with extremities mucronate or acerate, where there is greater concentration of microspines. A single category, ranged 156–307.7–413/3–7.1–11 µm, with stylote and strongylote modifications ( Fig. 3 View FIGURE 3 E–K). Trichodragmata occur in two categories, largest with 38–69–110 µm and the smallest with 12–19–29 µm, which the former more rare than the second ( Fig. 3 View FIGURE 3 L–M).
Ecology. The specimens were found from 20 to 30 m depth.
Geographical distribution. Northeastern coast of Brazil (Sergipe and Alagoas States).
Etymology. The specific epithet derives from the diagnostic characteristic of Myrmekioderma intrastrongyla sp. nov., the presence of strongyles only in the choanosome.
The new species belongs to the genus Myrmekioderma by the presence of trichodragmata, ectosomal skeleton formed by acanthoxeas perpendicular to the surface and a halichondroid choanosome. Its skeleton organization showed similarity with Didiscus Dendy, 1922 and Heteroxya Topsent, 1898 . However, the first differs from Myrmekioderma intrastrongyla sp. nov. by its discorhab-like microscleres and the latter by the absence of trichodragmata ( Hooper, 2002). Although in some species of Myrmekioderma , trichodragmata was also not found, Bergquist (1965) noted that in Heteroxya there is no mention of variability in spicule terminations and megascleres are extremely large (2000/35 µm).
The eight species of the genus have only oxeas as main spicular component, which can be modified to styles and strongyles (Table 2) ( Pulitzer-Finali, 1983; Hooper & Lévi, 1993). In Myrmekioderma intrastrongyla sp. nov., strongyles occur as other spicule category, larger than oxeas and distributed through the choanosome. Thus, only M. tuberculatum and M. intrastrongyla sp. nov. differ from others of the genus by the presence of these strongyles. However, in M. tuberculatum such spicules are rare and the trichodragmata are absent.
Despite Myrmekioderma being characterized by having more than one category of (acanth-)oxea ( Hooper, 2002), both M. tuberculatum as M. niveum have only a single category of oxeas. This can also be seen in M. intrastrongyla sp. nov., which differs from these species by the presence of an extra category of strongyles and trichodragmata, not found in both species ( Keller, 1891; Row, 1911; De Laubenfels, 1934).
Myrmekioderma granulatum has external morphology almost identical to paratypes of M. intrastrongyla sp. nov. Both species have massive form and surface composed by polygonal protuberances (honeycomb) separated by shallow and very characteristics grooves. However, M. granulatum differs from the new species by having two categories of (acanth-)oxea, a single category of trichodragmata and the absence of strongyles. Bergquist (1965) and Hooper et al. (1997) report a wide variation in the spicules morphology found in disjunct population of M. granulatum . Even raising the possibility of the existence of a species complex, Hooper et al. (1997) suggested that morphological data alone were insufficient to define the status of these populations. Despite this wide spicular variability, none of these populations showed choanosomal strongyles as found in M. intrastrongyla sp. nov.
In addition, the tubercular projections on the surface of Myrmekioderma rea observed by Diaz et al. (1993) are also quite similar to the one described for the new species ( Fig. 2 View FIGURE 2 B). However, the presence of strongyles in Myrmekioderma intrastrongyla sp. nov. versus the presence of oxeas and styles in M. rea differentiate the two species.
De Laubenfels (1934) described Myrmekioderma rea as Anacanthea , without characterizing adequately their spicular types. In 1953, he described M. styx mentioning the presence of two categories of (acanth-)oxeas and the presence of trichodragmata. Castellanos et al. (2003) synonymized M. styx sensu De Laubenfels (1953) as M. rea , while the remaining records of M. styx by other Caribbean authors were synonymized as Myrmekioderma gyroderma . They argued that the two species were different mainly by M. rea having oxeas and styles while M. gyroderma has only stout oxeas. However, the discussion of Castellanos et al. (2003) about this synonymy was scarce, since the change should be explained and demonstrated by comparing the characteristics of type material. This fact will be fixed by Valderrama & Zea that will publish a paper with the comparison of holotypes of both species. Nevertheless, the combination of external morphology, presence of strongyles and two types of trichodragmata, make Myrmekioderma intrastrongyla sp. nov. distinct from any other Myrmekioderma described for the Western Atlantic.
Both prior records of Myrmekioderma from Brazil need a reevaluation of the specimens to define their precise identity. When Mothes et al. (2004) recorded M. rea from Brazil, they reported that the only difference from Caribbean specimens is the lack of trichodragmata. However, we realize other important differences, such as the variation in the size of spicules in their respective categories, and the absence of styles in the Maranhão material, versus the common occurrence of styles in the Caribbean material. Thus, it is possible that the material examined by Mothes et al. (2004) is more closely related to M. gyroderma , since it has greater oxeas (Table 2) with few modifications, and no trichodragmata, as described by Alcolado (1984). Furthermore, Mothes et al. (2004) synonymized the record of M. styx made by De Rosa– Barbosa (1995) for the Rio Grande do Sul State as M. rea . The presence of oxeas and styles, smaller acanthoxeas and only one category of trichodragmata resemble what has been described by Diaz et al. (1993) to M. rea . However, we note that in the description of the Brazilian material the megascleres have dimensions much larger than any other Myrmekioderma previously described (Table 2). Thus, while we consider it necessary to reassess these specimens to establish their precise identities, Myrmekioderma intrastrongyla sp. nov. does not resemble any of them as it has strongyles as part of its complement of megascleres, as well as two categories of trichodragmata.
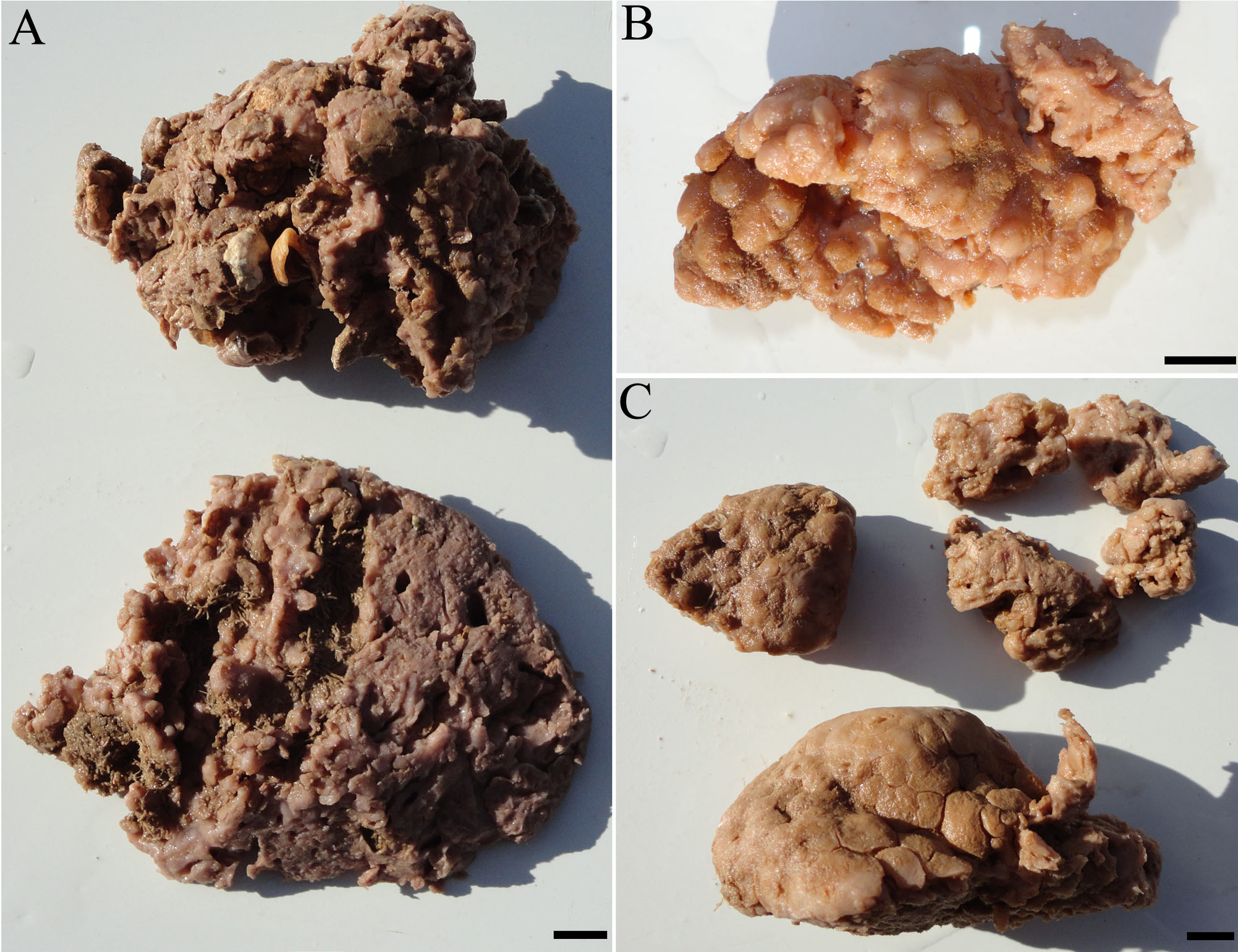

TABLE 1. Data micrometric of specimens types of Myrmekioderma intrastrongyla sp. nov. Values are in micrometres (µm), expressed as follow: minimum – mean – maximum; length / width.
| Specimens | Strongyles Acanthoxeas | Trichodragmata I | Trichodragmata II |
|---|---|---|---|
| MNRJ16784 (Holotype) | 312–554.5–675/6–10.8–12 287–349.5–412/6–7.6–10 | 38–73.1–89 | 12–16.5–28 |
| UFPEPOR1528 | 300–505.3–670/10–12.8–20 270–317.3–360/4–7.5–11 | 38–61.7–128 | 11–20–29 |
| UFSPOR184 | 412–538–725/8–10.5–13 212–264.4–312/4–7.2–10 | 38–59.7–90 | 12–15.7–22 |
| UFPEPOR1529 | 325–491.6–662/6–10.3–13 195–241.1–279/6–6.6–9 | 38–51.9–80 | 12–18.4–29 |
| UFPEPOR1530 | 312–514.1–700/8–11–13 212–306.6–350/4–7–10 | 54–73.4–83 | 12–20.9–28 |
| UFSPOR188 | 412–551.2–737/6–9.7–11 237–323.7–326/6–7.2–10 | 48–73.6–109 | 12–20–28 |
| UFSPOR137 | 300–625.4–775/8–11.8–13 243–313–336/4–7–9 | 38–74.5–94 | 10–20–29 |
| UFSPOR160 | 324–621.2–792/9–11.5–15 156–346–408/3–6.7–9 | 38–64.5–85 | 11–22–29 |
| Discussion |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Myrmekioderma intrastrongyla
| Sandes, Joana & Pinheiro, Ulisses 2013 |
M. styx sensu
| De Laubenfels 1953 |