
Mycale (Naviculina) obscura ( Carter, 1882 )
|
publication ID |
https://doi.org/10.11646/zootaxa.4912.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:9536C1CF-4AEF-47F8-959B-48CD7A5392D8 |
|
DOI |
https://doi.org/10.5281/zenodo.4473096 |
|
persistent identifier |
https://treatment.plazi.org/id/361087A7-FF59-FF00-55AB-F966532ECCB9 |
|
treatment provided by |
Plazi |
|
scientific name |
Mycale (Naviculina) obscura ( Carter, 1882 ) |
| status |
|
Mycale (Naviculina) obscura ( Carter, 1882) View in CoL
Figs 102 View FIGURE 102 a–f, 103a–i
Esperia obscura Carter, 1882: 299 View in CoL , pl. XI fig. 18.
Mycale obscura View in CoL ; Hentschel 1911: 302, fig. 9.
Mycale (Aegogropila) obscura View in CoL ; Shaw, 1927: 424 (no description).
Mycale (Naviculina) obscura View in CoL ; Hajdu 1999: 228.
Material examined. ZMA Por. 01602, Indonesia, Nusa Tenggara, between Komodo and Sumbawa, 8.5°S 119.125°E GoogleMaps , dead corals and sand, depth 73 m, dredge, coll. Siboga expedition stat. 310, field nr. SE110 VI, 12 February 1900 ; ZMA Por. 02888, Timor Leste, anchorage between Nusa Besi and NE point of Timor , 8.42°S 127.3066°E, sand and coral, depth 27–54 m, dredge and trawl, coll. Siboga Expedition stat. 282, field nr. SE675 GoogleMaps II, 15 January 1900 ; ZMA Por. 02902, Philippines, Sulu Archipelago, anchorage off North Ubian , 6.125°N 120.4333°E, lithothamnion bottom, depth 16–23 m, dredge, coll. Siboga Expedition stat. 099, field nr. SE 1474XI, 28 June 1899 GoogleMaps ; ZMA Por. 02903, Philippines, Sulu Archipelago, anchorage off North Ubian , 6.125°N 120.4333°E, lithothamnion bottom, depth 16–23 m, dredge, coll. Siboga Expedition stat. 099, field nr. SE 1474VIII, 28 June 1899 GoogleMaps ; ZMA Por. 08912, Indonesia, Sulawesi, SE Sulawesi, North East Take Bone Rate, south of Tarupa Kecil , 6.5°S 121.1333, depth 10–15 m , SCUBA, coll R. W.M. van Soest, Indonesian-Dutch Snellius II Expedition, stat. 139, field nr 139/ IV/31 , 25 September 1984 (live color pink) ; ZMA Por. 11067, Palau Islands , coll. M.K. Harper, field nr. 95–049, no further data (orange) ; ZMA Por. 11068, Palau Islands , coll. M.K. Harper, field nr. NCI2096, no further data (orange) ; ZMA Por. 12040, Seychelles, Amirantes, northern slope of Île Desnoeufs Platform , 6.2167°S 53.0167°E, depth 12–15 m GoogleMaps , SCUBA, coll. R. W.M. van Soest, Netherlands Indian Ocean Expedition stat.764, field nr IOP-E 783/05, 2 January 1993 (orange-yellow) ; ZMA Por. 13274, Indonesia, Sulawesi, SW Sulawesi, Kudingareng Keke , depth 15 m , SCUBA, coll. N.J. de Voogd, field nr. KK/NV/070497/01, 7 April 1997 (bright orange) ; ZMA Por. 13705, Seychelles, Mahé, NW coast, Beauvallon Bay, Vista do Mar , 4.5667°S 55.4333°E, depth 0.5–9 m, snorkeling, coll GoogleMaps . R. W.M. van Soest, Netherlands Indian Ocean Expedition stat.609, field nr. IOP-E 609/04, 11 December 1992 ; ZMA Por. 13894, Oman, Masirah Island, Ras Hilf , depth 13 m , SCUBA, coll. R.G. Moolenbeek, 15 November 1998 (brownish bordeaux-red).
Description ( Figs 102 View FIGURE 102 a–d). Thinly to massively encrusting on dead coral rubble, with an irregularly outlined, folded-lobate shape in larger individuals ( Fig. 102a View FIGURE 102 ), more compact in smaller specimens ( Figs 102 View FIGURE 102 b–d) (no in situ images were available to us). Surface predominantly smooth, semi-transparent, with detachable surface skeleton, distinctly reticulated in life. Size of individual specimens and fragments variable, up to 8 x 3 x cm. Color in life ranging from bright orange to pinkish red-brown, in preservation transparent dirty white to reddish and brownish. Consistency soft.
Skeleton ( Figs 102 View FIGURE 102 e–f). The choanosome ( Fig. 102f View FIGURE 102 ) is traversed by strongly developed spicule tracts, up to 150 µm in thickness (up to 25 spicules in cross section), which divide into thinner tracts and fan out just below the surface to carry the tangential ectosomal skeleton. The surface reticulation ( Fig. 102e View FIGURE 102 ) consists of intercrossing tracts of 15–35 µm diameter making meshes of 120–250 µm in widest size. Rosettes of anisochelae I, 90–110 µm in diameter, are scattered in the surface region ( Fig. 102e View FIGURE 102 ), often near or on the crossings of the spicule tracts. Other microscleres are found throughout the ectosome and choanosome.
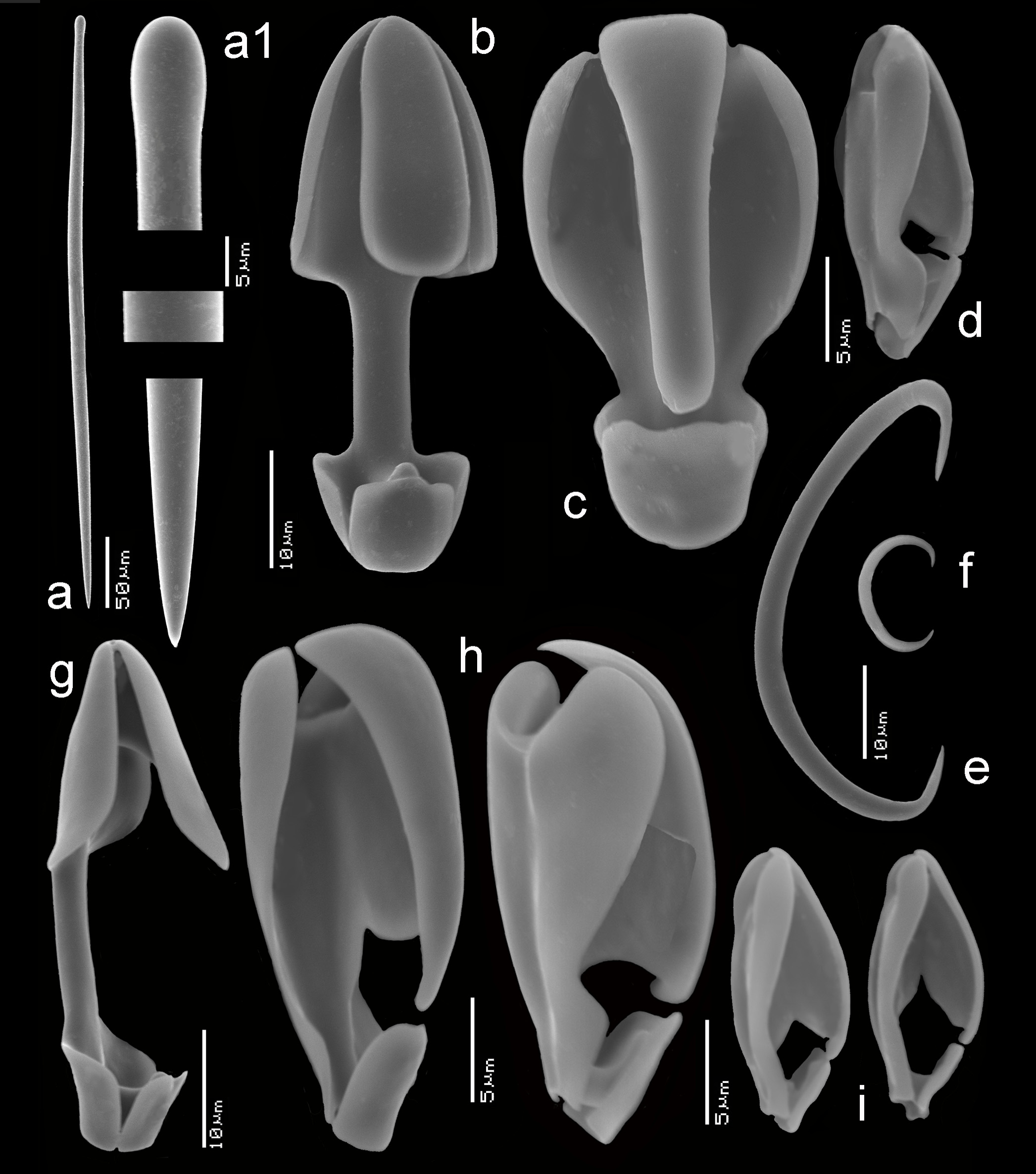
Spicules ( Figs 103 View FIGURE 103 a–i). Mycalostyles, three categories of anisochelae, two categories of sigmas.
Mycalostyles ( Figs 103a,a View FIGURE 103 1 View FIGURE 1 ), robust, usually straight, with well-developed heads and subterminal constriction, 312– 419.5 –481 x 6– 7.9 – 11 µm.
Anisochelae I ( Figs 103b,g View FIGURE 103 ), ‘normal’ shaped, with well-developed alae, upper median alae extended outwards, lower median alae often with small upward projection, free part of shaft 25–35% of spicule length, quite variable over the wide range of occurrence, 27– 40.9 – 54 µm.
Anisochelae II ( Figs 103b,h View FIGURE 103 ), naviculichelae, robust, with rounded outline, both sides slightly different in shape ( Figs 103h View FIGURE 103 ), variable with regard of closure of the central plate and merger of the median upper and lower alae, variable in length over the range of occurrence, 18– 24.3 – 32 µm.
Anisochelae III ( Figs 103d,i View FIGURE 103 ), naviculichelae, oval in outline, usually the central plate is entirely closing off the space between the upper and lower alae, variable in length over the range of occurrence, 9– 17.8 – 21 µm.
Sigmas I ( Fig. 103e View FIGURE 103 ), occasionally rarely present, robust, up to 3.5 µm in thickness, asymmetrical, comparatively shallow-curved, 39– 59.8 – 82 µm.
Sigmas II ( Fig. 103f View FIGURE 103 ), thin, 9– 12.3 – 22 µm.
Distribution and ecology. Indonesia, Timor Leste, Philippines, Palau, Seychelles, Oman, Western Australia, Tasmania, from shallow-water reef flat down to sandy bottoms at 73 m.
Remarks. The species is set off against closely related M (N.) cleistochela by its lack of flagellated sigmas Ib and toxas. M. (N.) cliftoni and M. (N.) microxea differ in the absence of ‘normal’ anisochelae.
Spicule size data show an unusually large range in this species and also presence and abundance of various microscleres show comparatively large fluctuation among the specimens. Possibly there are regional trends in these features.
The present species is close to lobate Korean Mycale (Naviculina) chungae Lerner & Hajdu, 2002 (originally as Mycale (Aegogropila) hentscheli Sim & Lee, 2001: 27 , figs 2A–M, figs 3A–F). The only significant difference is the apparent absence of sigmas II. Brazilian M. (N.) arcuiris Lerner & Hajdu, 2002 is also quite similar to the present species, and they even have the two sigma sizes. Geographic separation is the major factor separating this species from the present one.
Outside our study area, two additional Mycale (Naviculina) species similar to M. (N.) obscura have been described from Korea by Sim & Kang 2004. M. (N.) ulleungensis has two size categories of normal anisochelae and only a single category of naviculichelae and sigmas, whereas M. (N.) neunggulensis has one size category of normal anisochelae, a single category of naviculichelae and two size categories of sigmas. Both are thus clearly different from M. (N.) obscura .
Additional Mycale (Naviculina) species from the region


| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
| VI |
Mykotektet, National Veterinary Institute |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Mycale |
Mycale (Naviculina) obscura ( Carter, 1882 )
| Van, Rob W. M., Aryasari, Ratih & De, Nicole J. 2021 |
Mycale (Naviculina) obscura
| Hajdu, E. 1999: 228 |
Mycale (Aegogropila) obscura
| Shaw, M. E. 1927: 424 |
Mycale obscura
| Hentschel, E. 1911: 302 |
Esperia obscura
| Carter, H. J. 1882: 299 |