
Alligatorellus
|
publication ID |
https://doi.org/ 10.1111/zoj.12400 |
|
persistent identifier |
https://treatment.plazi.org/id/F75787E0-FF9B-FFC8-0251-042180C7BE2E |
|
treatment provided by |
Marcus |
|
scientific name |
Alligatorellus |
| status |
|
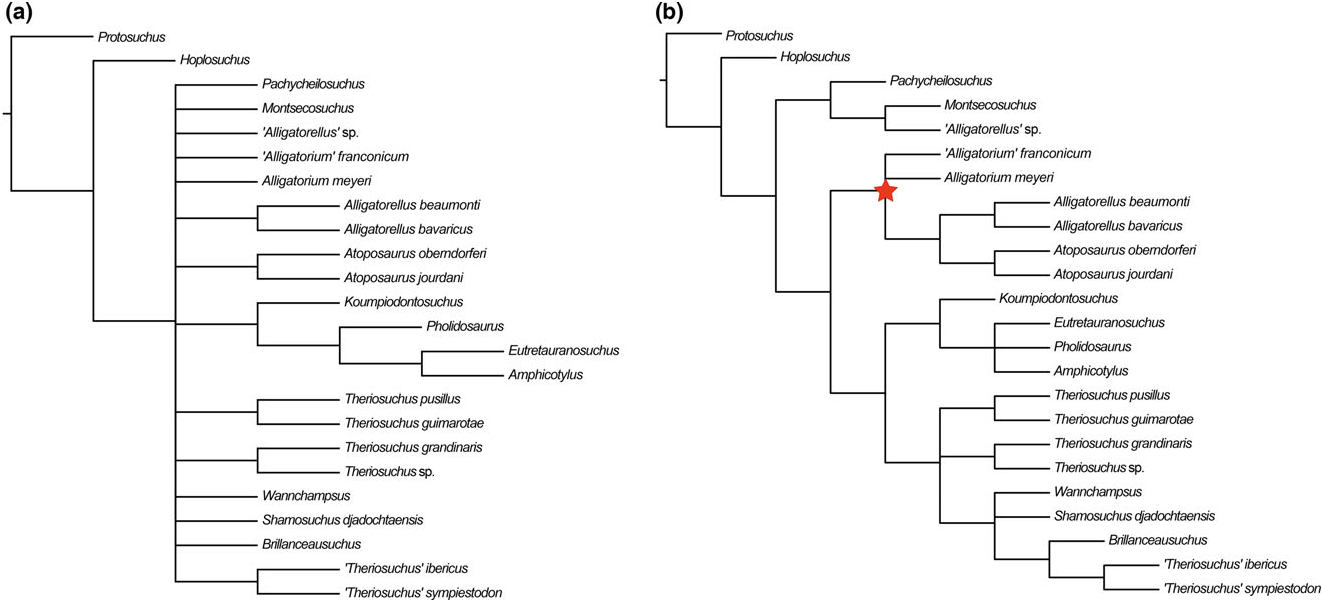
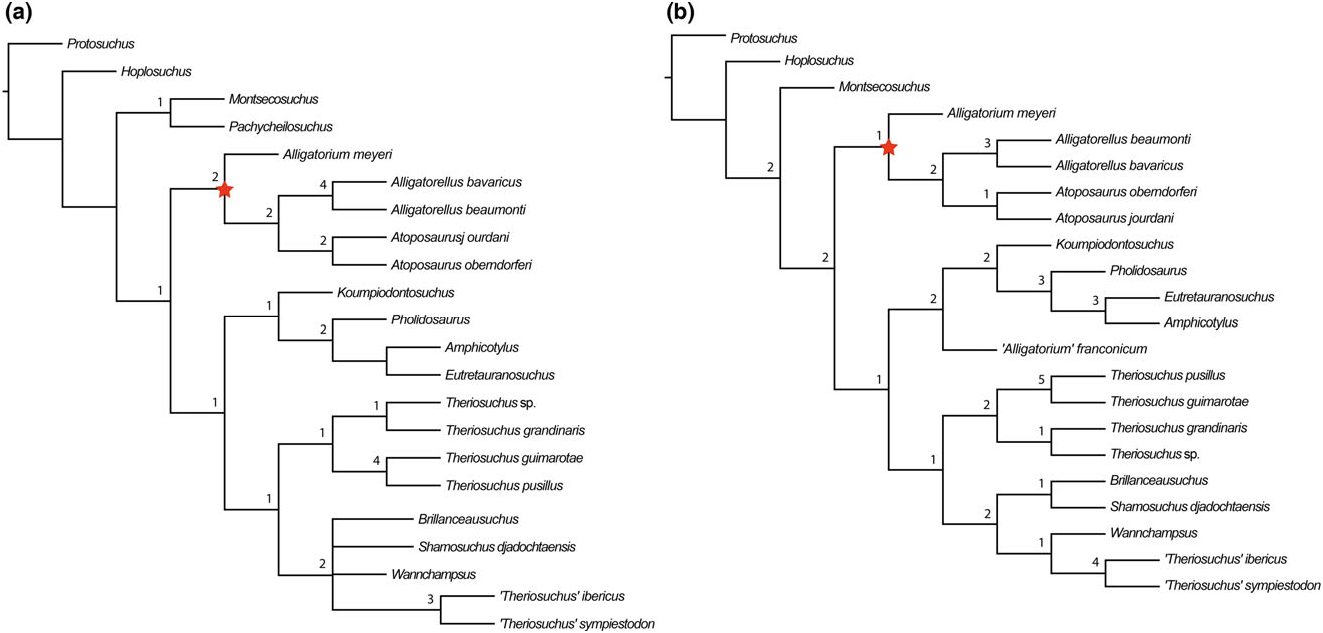
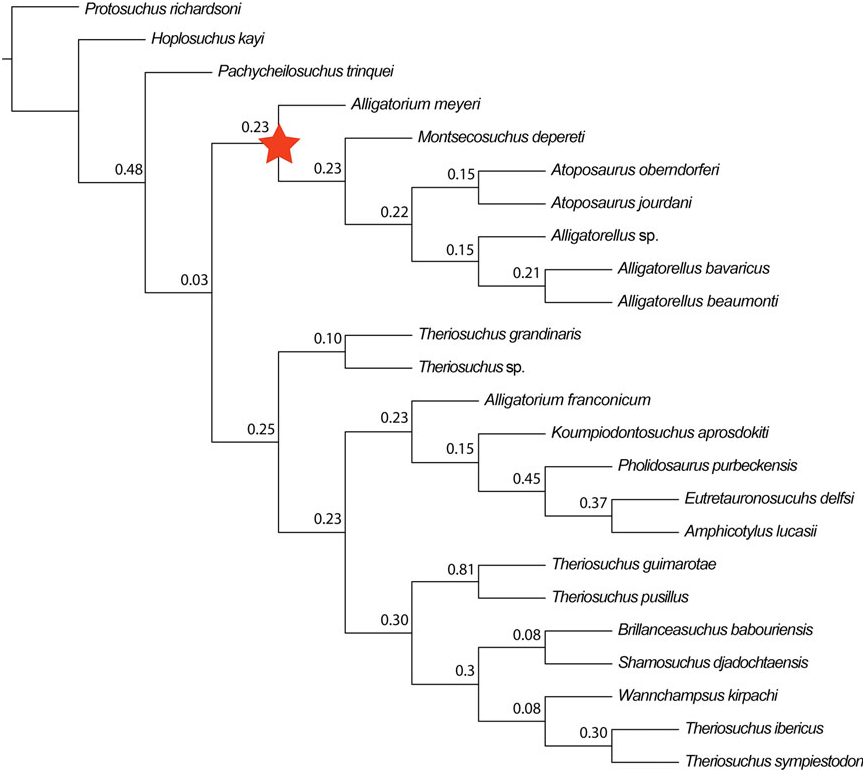
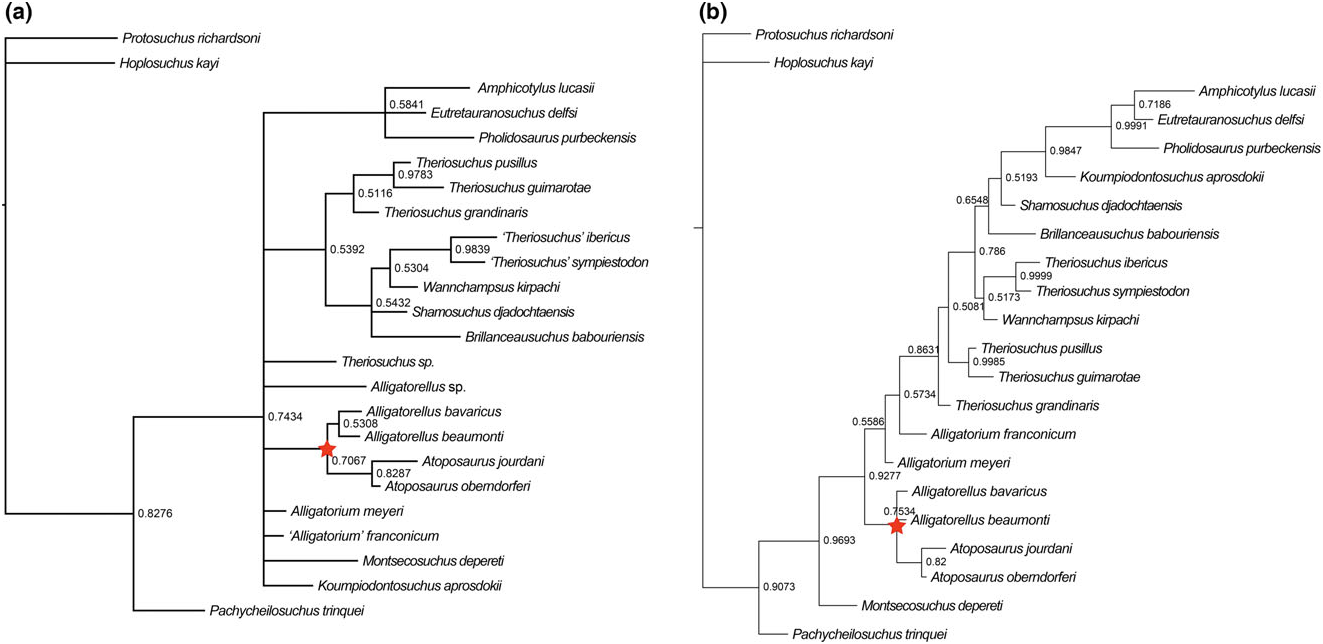
Alligatorellus is united with Atoposaurus within all of our analyses ( Figs 4 – 7 View Figure 4 View Figure 5 View Figure 6 View Figure 7 ). This is based on a range of character states, including: (1) a slit-like (i.e. mediolaterally narrow and anteroposteriorly elongated) external supratemporal fenestra (not visible in Atoposaurus oberndorferi owing to preservation) (C17.2); (2) smooth lateral surface of anterior jugal process near maxillary contact, not stippled or striated (C51; note that we did not code this as a separate character state owing to potential duplication, as in Atoposaurus , the entire external surface of the skull is smooth and unsculpted); (3) straight ventral edge of maxilla in lateral view (C52.0), similar to protosuchians (not sinusoidal or convex as in other neosuchians); (4) minimum mediolateral width between supratemporal fenestrae more than one-third of total width of cranial table (C126.1), acquired in parallel with Montsecosuchus , Brillanceausuchus , T. pusillus , and ‘ T.’ sympiestodon ; and (5) postorbital bar between orbit and supratemporal fenestra very narrow (with respect to lateral edge of postorbital lateral to supratemporal fenestra) and unsculpted, with superficial furrow on dorsal surface of postorbital connecting anterior edge of supratemporal fenestra to the posterior edge of and orbital (C128.2).
Included species
Atoposaurus jourdani (type species, named first in von Meyer, 1850) and Atoposaurus oberndorferi .
Distribution
Late Jurassic of southern France and south-east Germany.
Previous diagnoses and comments
Both species of Atoposaurus were named by von Meyer (1850), with Atoposaurus jourdani receiving a full description by von Meyer (1851). Wellnhofer (1971) was the first to present a diagnosis for Atoposaurus based on the specimens from France and Germany, noting the lack of dermal armour, a feature that could be related to either ontogeny or taphonomy (Schwarz & Salisbury, 2005). Subsequently, Atoposaurus and its constituent species have largely been considered to be nomina dubia, and often regarded as juvenile representatives of Alligatorellus and/or Alligatorium (e.g. Clark, 1986; Buscalioni & Sanz, 1988). Clark (1986) considered Alligatorellus , Alligatorium , and Atoposaurus to be the same taxon represented by different growth stages. If this were the case, then Atoposaurus von Meyer, 1850 ; would retain priority, and Alligatorium Jourdan, 1862 , along with Alligatorellus Gervais, 1871 , would be synonymized with Atoposaurus . Almost all subsequent phylogenetic analyses have included just Alligatorium based on this conclusion, without consideration of the other taxa. The present analysis is the first to consider both potential species of Atoposaurus as independent OTUs, and finds them to be sister taxa in all cases ( Figs 4 – 7 View Figure 4 View Figure 5 View Figure 6 View Figure 7 ), thus supporting their generic assignment.
Despite noting the same features in Alligatorellus, Wellnhofer (1971; see also Steel, 1973) stated that the presence of large orbits, a closed internal supratemporal fenestra, and divided external nares were all features defining Atoposaurus . The presence of an inwardly displaced postorbital bar (Steel, 1973) is not clear owing to the preservation of available specimens of Atoposaurus oberndorferi , but does appear to be a feature of Atoposaurus jourdani . Four of the five synapomorphies that we identify for Atoposaurus (S1 – 3, S5) are contentious as they could be indicative of a juvenile phase of growth (e.g. Joffe, 1967), but equally probably they could represent the retention of juvenile characteristics through paedomorphism related to the relatively small body size of Atoposaurus . Unfortunately, based on currently available specimens, it is impossible to distinguish between these two hypotheses (Tennant & Mannion, 2014).
Revised diagnosis and discussion
(S1) Dorsal cranial bones comprising the skull roof unsculpted (C1.0): This lack of dermal sculpting, combined with their overall diminutive size, indicates that Atoposaurus specimens might be represented by juveniles. However, it cannot be definitively confirmed that Atoposaurus is a juvenile representative of other contemporaneous atoposaurids based on allometric growth patterns alone (Tennant & Mannion, 2014), and it is likely that Atoposaurus , Alligatorellus , and Alligatorium represent three distinct genera, as our results indicate ( Figs 4 – 7 View Figure 4 View Figure 5 View Figure 6 View Figure 7 ).
(S2) External surface of snout unsculpted (C3.0): We consider this to be a distinct feature from S1, as in Alligatorellus there is a different pattern of sculpting between the cranial table and the rostrum. Atoposaurus is similar to Alligatorellus in this respect, completely lacking any evidence of cranial ornamentation, although this cannot be assessed properly in Atoposaurus oberndorferi owing to the mode of preservation of the holotype specimen. As with S1, this character is likely to be highly influenced by either ontogeny or paedomorphism (Joffe, 1967).
(S3) Skull anteroposterior length to orbit length ratio <3.0 (C27.0): This feature is unique to Atoposaurus , and represents the characteristic proportionally large orbits and short snout of this taxon, noted by Joffe (1967) to be indicative of a juvenile status. Other atoposaurids have a ratio of between 3.0 and 4.0. Karatausuchus has a ratio of 3.36, slightly higher than Alligatorellus bavaricus (3.12), which approaches the state boundary for Atoposaurus , but it is likely that Karatausuchus represents a juvenile specimen of a (probably non-atoposaurid) crocodyliform (Storrs & Efimov, 2000; see below).
(S4) 50 or more caudal vertebrae (C276.1): Complete axial columns are rarely preserved in specimens previously assigned to Atoposauridae , and the proportional numbers of cervical, dorsal, sacral, and caudal vertebrae remain poorly known, especially for Theriosuchus . Both species of Alligatorellus preserve complete and articulated caudal vertebral series, and have 40 vertebrae each. Montsecosuchus appears to only have 21 caudal vertebrae, and Pachycheilosuchus has just 18 (Rogers, 2003). Protosuchus richardsoni has 39 caudal vertebrae, and Karatausuchus has 46 (Storrs & Efimov, 2000), approaching the number for Atoposaurus , but no other crocodyliform taxon has 50 vertebrae. The presence of 50 or more caudal vertebrae, in all specimens of Atoposaurus in which this feature can be measured, is not known in any other mesoeucrocodylian taxon, and cannot be explained by ontogeny (Tennant & Mannion, 2014); therefore, we regard it as a diagnostic feature for Atoposaurus , irrespective of the ontogenetic stage of the specimens, and therefore consider Atoposaurus to be a valid taxon.
(S5) Osteoderms absent: The lack of osteoderms is unlikely to be a taphonomic artefact (contra Schwarz-Wings et al., 2011), and is either a feature associated with extreme dwarfism in Atoposaurus , or relates to their lack of development in juvenile individuals. The only other putative atoposaurid that is similar in this respect is Karatausuchus , which Storrs & Efimov (2000) described as having reduced dermal ossicles.
ATOPOSAURUS JOURDANI VON MEYER, 1850 (TYPE SPECIES)
Type locality and horizon
Unknown bed, Kimmeridgian (Late Jurassic); Cerin, Ain, France.
Type specimen
MHNL 15679 View Materials , articulated partial skeleton comprising dorsally flattened skeleton and skull, missing the posterior-most caudal vertebrae, with distal hindlimbs and distal left forearm preserved as impressions .
Referred specimen
MHNL 15680 View Materials (same locality as type specimen), posterior half of articulated skeleton, including trunk vertebrae and forearms .
Previous diagnoses and comments
von Meyer (1851) named Atoposaurus jourdani , and described this taxon in a subsequent paper (von Meyer, 1851). We find that a unique combination of metric characters, almost exclusively regarding the relative proportions of the forelimb and hindlimb elements, can be used to distinguish this taxon from Atoposaurus oberndorferi , along with a single autapomorphy.
Revised diagnosis and discussion
(S1) Skull mediolateral width to orbit width ratio of 1.80 (C28.0): This represents the lowest ratio for all atoposaurids, demonstrating that the orbits comprise almost the entire mediolateral width of the skull, separated by the narrow frontals. This is similar to Alligatorellus bavaricus , which also has enlarged orbits, but slightly mediolaterally wider frontals between the orbits (Tennant & Mannion, 2014).
(S2*) Six cervical vertebrae (C266.0): This character state is unique amongst all putative atoposaurids, with all others possessing seven cervical vertebrae. We were unable to determine this character state for any specimens assigned to Theriosuchus owing to their preservation and/ or incompleteness. This cervical count is distinct from Protosuchus richardsoni (nine) and Hoplosuchus (11), as well as Karatausuchus (eight; Storrs & Efimov, 2000) ( Table 2).
(S3) Forelimb length to hindlimb length ratio of 0.63 (C285.0): This character state is similar to Protosuchus richardsoni (0.65) and Montsecosuchus (0.63). Atoposaurus jourdani is distinct in this respect from other atoposaurids, including Alligatorellus beaumonti , which has a ratio of 0.81, and Atoposaurus oberndorferi , which has a ratio of 0.78. Alligatorellus bavaricus has an intermediate ratio of 0.76 ( Table 3).
(S4) Humerus length to femur length ratio of 0.67: This feature is similar to Protosuchus richardsoni (0.66) and Montsecosuchus (0.70). Alligatorellus beaumonti is similar, with a ratio of 0.75, but Alligatorellus bavaricus is quite distinct, with a ratio of 0.89, similar to Atoposaurus oberndorferi (0.89). Karatausuchus also has similar limb proportions, with a ratio of 0.73. Theriosuchus pusillus falls within the range for atoposaurids, with a ratio of 0.76, but T. guimarotae is distinct with a ratio of 0.98, approaching that for Pachycheilosuchus (1.09) and Brillanceausuchus (1.11) ( Table 3).
(S5) Radius to tibia length of 0.61 (C286.0): This character state is similar to Protosuchus richardsoni (0.63) and Montsecosuchus (0.64), along with Pachycheilosuchus (0.64). Theriosuchus pusillus has a proportionally long radius to tibia ratio (0.55), with this value only being exceeded by Karatausuchus (0.47). Based on Wellnhofer (1971), ‘ Alligatorium ’ franconicum has the most extreme value, with a ratio of 0.89, reflecting a proportionally long radius. This value is similar to Brillanceausuchus (0.88). Atoposaurus oberndorferi and Alligatorium meyeri each have a ratio of 0.74, similar to Shamosuchus (0.72) and Hoplosuchus (0.76) ( Table 3).
(S6) Metatarsals longitudinally grooved (C302.0): This feature also characterizes Alligatorellus beaumonti and Montsecosuchus (Tennant & Mannion, 2014) , in contrast to the smooth and flat metatarsals that characterize most other mesoeucrocodylians. However, we are cautious in our interpretation of this feature, as there remains the possibility that it could represent post-mortem crushing of the delicate long bones in the tarsus.
ATOPOSAURUS OBERNDORFERI VON MEYER, 1850
Type locality and horizon
Solnhofen Formation, early Tithonian (Late Jurassic, Hybonoticeras hybonotum ammonoid zone); Kelheim, Eichst € att, Bavaria, Germany .
Type specimen
TM 3956 , near-complete skeleton, missing only the dorsal part of the skull and posterior portion of the tail .
Referred specimen
BSPG 1901 I 12, a counterpart specimen of a different individual comprising the impression of the complete skull and skeleton in lateral view.
Previous diagnoses and comments
Wellnhofer (1971) diagnosed Atoposaurus oberndorferi primarily on several size-based characteristics, but these are unlikely to represent diagnostic morphological characters. He also noted the presence of five premaxillary and eight maxillary teeth, but this could not be confirmed via observation of the type specimen because of the way in which it is preserved, and was not illustrated in the figure of the referred specimen in Wellnhofer (1971). Steel (1973) followed the diagnosis of Wellnhofer (1971), and also suggested that the inwardly displaced postorbital bar was diagnostic of Atoposaurus oberndorferi ; however, this feature is now recognized as characterizing Atoposauridae (see above). Furthermore, because of the lateral compression of the type specimen, it was not possible to directly confirm the presence of this feature in Atoposaurus oberndorferi , and it is not figured by Wellnhofer (1971), and therefore cannot be supported. We present a revised diagnosis based on examination of the type specimen for Atoposaurus oberndorferi , and tentatively consider it to be a valid taxon based on three ambiguous autapomorphies.
Revised diagnosis and discussion
(S1) Skull anteroposterior length to width ratio ~2.00 (1.98) (C25.0): This feature is tentatively considered to be diagnostic for Atoposaurus oberndorferi , as the skull is highly incomplete and preserved only in ventrolateral aspect. This estimated skull length-to-width ratio is high, similar to Eutretauranosuchus (1.97), Montsecosuchus (1.80), which is represented by a mature specimen (Buscalioni & Sanz, 1990a), and Theriosuchus guimarotae (1.82) (Schwarz & Salisbury, 2005). Atoposaurus jourdani has a much lower ratio (1.28), more similar to Protosuchus (1.31) and Hoplosuchus (1.35). The only taxa that have higher ratios are Alligatorellus (2.06 – 2.21), Alligatorium meyeri (2.26), ‘ Alligatorium ’ franconicum (2.77), and Koumpiodontosuchus (2.04) ( Table 3).
(S2) Skull anteroposterior length to orbit length ratio <3.00 (2.86) (C27.0): As with (S1), we are cautious with our interpretation of this character state based on the way in which the observed type specimen of Atoposaurus oberndorferi is preserved, exposing the enlarged orbit only in ventral aspect. The only taxon with a lower ratio is Atoposaurus jourdani (2.33), with Alligatorellus and Alligatorium possessing ratio values between 3.0 and 4.0. Protosuchus has a ratio of 4.52 ( Table 3), an intermediate value between atoposaurids and higher neosuchians.
(S3) Inwardly (dorsally) displaced splenial on the ventral mandibular surface (C234.1): In all other taxa we analysed in which the ventral surface of the mandible was exposed, the anterior portion of the splenial is confluent ventrally with the posterior cavity that is formed from the two posteriorly divergent mandibular rami. In Atoposaurus oberndorferi , the splenial is slightly inset at its contact with the dentary, a feature shared only with T. pusillus . The ventral side of the skull and mandibular region is not preserved in Atoposaurus jourdani , and this character state might also be present in that taxon too. Therefore, we are cautious in our retention of Atoposaurus oberndorferi as a distinct, second species of Atoposaurus .
Included species
Alligatorellus beaumonti (Gervais, 1871) , Alligatorellus bavaricus (Wellnhofer, 1971; sensu Tennant & Mannion, 2014).
Distribution
Late Jurassic of southern France and south-east Germany.
Previous diagnoses and comments
Alligatorellus was diagnosed by Wellnhofer (1971) based on its overall size, the shape of its skull, and its relatively large orbits, features that are all more widespread amongst atoposaurids and other smallbodied neosuchians. Wellnhofer (1971) originally described two subspecies of Alligatorellus beaumonti , based on relative sizes and differences and geographical distribution. Most recently, Tennant & Mannion (2014) documented a number of distinguishing characters between Alligatorellus beaumonti beaumonti from France and Alligatorellus beaumonti bavaricus from Germany, and reranked the latter to its own species, Alligatorellus bavaricus . Several of the diagnostic synapomorphies for Alligatorellus , presented below, might be related to ontogenetic factors, such as the heterogeneity of the cranial sculpting and the closed internal supratemporal fenestra (Joffe, 1967). However, these features could also be related to the proposed ‘dwarfism’ for atoposaurids, and there are other indicators that the available specimens of Alligatorellus represent a reasonably mature state of growth, such as neurocentral fusion and the degree of fusion of the cranial bones (Joffe, 1967; Tennant & Mannion, 2014).
Schwarz-Wings et al. (2011) referred a partial skeleton, MB.R.3632, from the early Tithonian of Franconia, Germany ( Gravesia gigas ammonoid zone) to Alligatorellus sp. , but Tennant & Mannion (2014) concluded that this specimen could only be referred to as Atoposauridae indet. In most of our analyses, this specimen is recovered as an indeterminate non-atoposaurid taxon ( Figs 4 View Figure 4 , 7A View Figure 7 ). However, when we used implied weighting, this specimen groups with the other species of Alligatorellus within Atoposauridae ( Fig. 6 View Figure 6 ). Therefore, we tentatively regard its status as Alligatorellus sp. to be valid.
Revised diagnosis and discussion
(S1) Cranial table sculpting composed of homogeneous, subcircular shallow pits (C2.2): The cranial sculpting pattern for Alligatorellus is distinct from that of Atoposaurus , which has a smooth dorsal surface, and from Alligatorium and Theriosuchus in which the sculpting is much more prominent. It is similar to Wannchampsus , which is also lightly sculpted. The reduction or lack of sculpting has been noted in smaller specimens of the basal mesoeucrocodylian Zosuchus (Pol & Norell, 2004a,b), as well as the protosuchian Gobiosuchus (Osmolska, Hua & Buffetaut, 1997) .
(S2) Rostrum unsculpted or relatively less than the cranial table (C3.1): Similar to (S1), the sculpting of the rostrum is relatively light compared with Alligatorium meyeri , T. pusillus , and Wannchampsus . Distinct from these taxa, however, is how the degree of sculpting appears to decrease anteriorly for Alligatorellus , with a more prominent pattern on the cranial table, and almost no sculpting on the dorsal surface of the rostrum. This morphology is similar to that seen in the paratype of Isisfordia , which is represented by an adult specimen (Salisbury et al., 2006), and Pachycheilosuchus , which is known from mature individuals (Rogers, 2003). In other taxa represented by mature specimens, such as T. pusillus and Rugosuchus (Wu, Cheng & Russell, 2001a) , sculpting patterns are homogeneous across the entire dorsal surface of the skull. Alligatorium meyeri is unusual in that the degree of sculpting remains the same between the rostrum and cranial table, but anteriorly the subcircular pits become more elongated, a feature visible in the counterpart to its holotype specimen (MNHL 15462), and which helps to distinguish it from Alligatorellus .
(S3) Closed internal supratemporal fenestra (C16.1): This feature refers to the lack of opening of the internal supratemporal fenestra, as noted by Wellnhofer (1971). In all other specimens we observed, the internal supratemporal fenestra is completely open. Joffe (1967) described the opening as ‘slit-like’ for T. pusillus , and regarded it as indicative of an immature individual. However, our observations of the paratype specimen (NHMUK PV OR48330) did not confirm this, and the internal fenestrae appear to be fully open. Because of poor preservation, we were unable to determine whether the morphology of the internal supratemporal fenestra was open or closed in any specimen of Atoposaurus .
(S4) Frontal maximal mediolateral width between the orbits narrower than maximal width of nasals (C97.1): This character state relates to the proportionally large size of the orbits, which occupy the majority of the mediolateral width of the dorsal surface of the skull, with a proportionally narrow interorbital region composed of the fused frontals. Although this feature is shared by many other neosuchians, including Theriosuchus and Wannchampsus , the frontals are distinctly narrower in Alligatorellus . In protosuchians, such as Protosuchus and Hoplosuchus , the mediolateral width of the frontal is broader than the nasal, because in these taxa the orbit is more laterally facing, and therefore does not occupy as much of the mediolateral width of the skull in dorsal view.
(S5) Broad frontal anterior process with parallel lateral margins, not constricted (C109.0): This feature is distinct from the morphology described in (S4), and relates exclusively to the development of the frontals anteriorly to the anterior margin of the orbits, excluding the morphology of any frontal anterior process where present. The broad anterior edge of the frontal with parallel lateral edges in Alligatorellus is similar to paralligatorids, T. pusillus , and Montsecosuchus , but contrasts with T. guimarotae in which the mediolaterally constricted anterior portions of the frontals distinctly underlap the nasals (Schwarz & Salisbury, 2005).
(S6) Flat and unsculpted anterior supratemporal margins (C119.0): In Alligatorellus bavaricus , the supratemporal rims are flat and unsculpted along their entire medial edge, similar to protosuchians, Pachycheilosuchus , coelognathosuchians, Montsecosuchus , and Koumpiodontosuchus . However, in Alligatorellus beaumonti , there is a slight posterior development of the supratemporal margins, similar to Brillanceausuchus (specimen UP BBR 203). This is distinct from Alligatorium meyeri , Wannchampsus , and all species referred to Theriosuchus , in which the supratemporal rims are consistently well developed along their entire medial margin.
(S7*) Anterior process of squamosal extends to the posterior orbital margin (C144.0): This character state appears to be diagnostic for Alligatorellus , although we are cautious in this assignment, as the postorbital region is poorly preserved, and the exact morphology of the postorbital with respect to the other posterior periorbital elements is difficult to assess. However, in the holotypes of Alligatorellus beaumonti and Alligatorellus bavaricus , there is no notable suture on the dorsal surface of the skull table, lateral to the supratemporal fenestra, which would represent the suture between the posterior process of the postorbital and the anterior process of the squamosal. We therefore infer that the anterior process of the squamosal reached the posterior orbital margin.
(S8) Posterodorsal margin of parietals and squamosals completely covers posterodorsal occipital region, excluding the supraoccipital from the dorsal surface of the skull (C197.1): This feature was proposed by Tennant & Mannion (2014) to be autapomorphic for Alligatorellus , but also appears to be present in a range of neosuchian taxa (e.g. Acynodon adriaticus ; Delfino et al., 2008b) in which the supraoccipital is excluded from the posterodorsal surface of the skull roof. We therefore consider it to only be locally diagnostic for Alligatorellus . In other mesoeucrocodylians, such as Mahajangasuchus (Turner & Buckley, 2008) , the supraoccipital is broadly visible in the midline portion of the posterodorsal region of the skull, contacting the parietals.
(S9) Smooth mandibular external surface, lacking sculpting (C201.0): This feature is difficult to observe in Alligatorellus bavaricus owing to the dorsal flattening of the holotype specimen, although what is visible indicates that the mandible, much like the anterior portion of the skull, lacks any sculpting pattern, unlike the dorsal surface of the cranial table. This is distinct from Theriosuchus and Wannchampsus , in which the sculpting pattern on the external surface of the dentaries and posterior mandibular elements is similar to that of the dorsal surface of the skull.
(S10) Proximal end of the radiale ‘hatchet-shaped’ (C290.1): This feature also characterizes MB.R.3632, and was used to refer this specimen to Alligatorellus (Schwarz-Wings et al., 2011) . However, this morphology is also shared by Wannchampsus (Adams, 2014) , but is not known in Theriosuchus specimens, owing to lack of preservation of the radiales. In other atoposaurids, the proximal end of the radiale is more expanded equidimensionally, lacking the asymmetry observed in Alligatorellus .
(S11) Proportionally short metatarsal I relative to metatarsals II–IV (C303.1): In other atoposaurids, metatarsals I – IV are almost equidimensional, possibly reflecting different locomotor adaptations in Alligatorellus .
(S12*)Dorsal surface of dorsal osteoderms completely sculpted (C304.0), with parallel and straight anterior and posterior margins (C308.1), and a longitudinal ridge along entire lateral margin (C311.1 and C312.1): The utility of osteoderms in atoposaurid systematics, particularly regarding Alligatorellus , was discussed by Tennant & Mannion (2014). These authors noted that the mediolateral position and anteroposterior extent of the dorsal keel, and its serial variation along the axial column, are diagnostic for Alligatorellus , as well as for other crocodyliforms (e.g. teleosauroids, eusuchians) that preserve a dorsal series of paravertebral osteoderms.
(S13) Caudal osteoderms with smooth, nonserrated edges (C327.1): This morphology is similar to the osteoderms preserved for T. guimarotae and T. pusillus , but distinct from Brillanceausuchus and Montsecosuchus in which the margins of the caudal osteoderms are serrated. Serrated edges might also be present in caudal osteoderms of MB.R.3632, based on at least three caudal osteoderms disassociated from the main osteoderm shield preserved on the specimen slab. However, we cannot discount the possibility that these elements are accessory dorsal osteoderms, as found in Montsecosuchus and in the proximal caudal series of Alligatorellus beaumonti .
ALLIGATORELLUS BEAUMONTI GERVAIS, 1871 (TYPE SPECIES) ALLIGATORELLUS BEAUMONTI BEAUMONTI WELLNHOFER, 1971
Type locality and horizon
Unknown bed, Kimmeridgian (Late Jurassic); Cerin, Ain, France.
Type specimen
MNHL 15639 , part and counterpart slabs comprising a near-complete and articulated skeleton, missing the distal-most caudal vertebrae (preserved as impressions) and part of the left forelimb. Parts of the skull roof and a large portion of the right maxilla, along with several axial fragments, are embedded into the counterpart slab .
Referred specimen
MNHL 15638, part slab comprising a near-complete and articulated skull and skeleton, missing just the distal-most caudal vertebrae, the right forelimb, and the distal left forelimb, all of which are preserved as impressions. The skull is exposed in ventrolateral aspect.
Previous diagnoses and comments
Alligatorellus beaumonti was originally named by Gervais (1871) for two specimens from the Late Jurassic of Cerin, eastern France. Subsequently, Wellnhofer (1971) diagnosed these specimens as a distinct subspecies, Alligatorellus beaumonti beaumonti . This was based largely on size differences between these and coeval specimens from Eichst att €, south-east Germany, for which Wellnhofer (1971) erected the subspecies Alligatorellus beaumonti bavaricus (see below). Together these specimens have largely been regarded as representing a single taxon, Alligatorellus beaumonti , by subsequent workers (e.g. Buscalioni & Sanz, 1988; Schwarz-Wings et al., 2011). However, Tennant & Mannion (2014) redescribed the German remains, and observed a number of morphological differences with the French material. They provided a revised diagnosis for Alligatorellus beaumonti , and re-ranked the German material as Alligatorellus bavaricus .
Wellnhofer (1971) noted that as in Theriosuchus , the external nares in Alligatorellus beaumonti are divided by an anterior projection of the nasals, a feature that also appears to be shared by Alligatorium meyeri and possibly Alligatorellus bavaricus , although the anterior-most portion of the snout in the holotype of the latter is damaged. Alligatorellus beaumonti is similar to Alligatorium meyeri in the presence of an unsculpted posterolateral ‘lobe’ of the squamosal, differing from Alligatorellus bavaricus in which the posterolateral corner of the squamosal instead displays orthogonal posterior and lateral edges. Buscalioni & Sanz (1988) suggested that another distinguishing feature between Alligatorellus beaumonti and Alligatorium meyeri is the contribution of the frontal to the supratemporal fenestra in the former; however, this feature is clearly also present in Alligatorium meyeri , and therefore cannot be used to distinguish the two taxa. The presence of a biserial osteoderm shield comprising singular sculpted osteoderms is not diagnostic for Alligatorellus (contra Wellnhofer, 1971), as it also characterizes both T. pusillus and Alligatorium meyeri . Tennant & Mannion (2014) proposed that the frontal width between the orbits being mediolaterally narrower than the nasals is an autapomorphy of Alligatorellus beaumonti ; however, this condition is not considered to be diagnostic here, as it is also known in a wide range of neosuchian taxa, and the width of the paired nasals in Alligatorellus bavaricus might have been underestimated. Alligatorellus beaumonti also has the reversed condition to Alligatorellus bavaricus , in that the anterior extension of the frontal exceeds the anterior margin of the orbits, similar to almost all other neosuchian taxa.
Revised diagnosis and discussion
(S1*) Frontal with unsculpted anterior and posterior portions, and sculpted medial surface: We elected not to code this as a distinct character state from that of S 3 in our matrix in order to avoid duplication of character states. Nonetheless, Tennant & Mannion (2014) identified this heterogeneity in sculpting pattern as distinct from other atoposaurids and Theriosuchus, and considered it to be autapomorphic of Alligatorellus beaumonti .
(S2) Surface of rostrum notably less sculpted than cranial table (C4.1): See S2 for Alligatorellus for discussion of this character state.
(S3) Relatively large lateral temporal fenestra, approximately 30% of the size of the orbit (C20.1): A lateral temporal fenestra of this size with respect to the orbit represents the intermediate condition in our analyses. This relatively large size is unique amongst atoposaurids, but is also shared with T. pusillus , Koumpiodontosuchus (Sweetman et al., 2015) , and protosuchians, as well as the advanced neosuchians Shamosuchus (Pol et al., 2009) , Isisfordia (Salisbury et al., 2006) , and Brillanceausuchus . In other taxa, such as Allodaposuchus precedens , the lateral temporal fenestra approaches the size of the orbit (Buscalioni et al., 2001).
(S4) Smooth contact between maxilla and jugal (C51.2): As noted above, the pattern of sculpting on the anterior portion of the dorsal surface of the skull is diagnostic for the different species of Alligatorellus . In Alligatorellus bavaricus , the entire dorsal surface is lightly sculpted, but Alligatorellus beaumonti has a smooth contact between the maxilla and jugal, similar to Atoposaurus and Hoplosuchus , although in both of these taxa the entire external surface of the skull is not ornamented. This is distinct from T. guimarotae and T. pusillus , which both have a contact in which the external surface is sculpted to the same degree as the rest of the cranial table, and from Brillanceausuchus , ‘ T. ’ ibericus , ‘ T. ’ sympiestodon , and Montsecosuchus , in which the contact is heavily striated.
(S5*) Medial longitudinal depression on posterior portion of nasal and anterior portion of frontal (C74.1): This is diagnostic of Alligatorellus beaumonti as a local autapomorphy, but is also present in the goniopholidid Amphicotylus (Mook, 1942) . This condition differs from that in Theriosuchus and a range of paralligatorids, including Brillanceausuchus , in which a distinct midline longitudinal crest develops.
(S6*) Posteromedial border of supratemporal fenestra forms a low sagittal rim (C119.1): This feature is considered to be locally autapomorphic, as it is also present in Brillanceausuchus . Alligatorellus bavaricus and Atoposaurus have no supratemporal rim development, and the rims are strongly developed along the whole medial edge of the external supratemporal fenestra in Alligatorium meyeri , T. pusillus , and Wannchampsus (Adams, 2014) .
(S7*) Smooth and unsculpted region on anterior portion of squamosal nearing orbit and posterolateral process of squamosal (C148.1): This feature appears to be locally diagnostic, but is also shared by Khoratosuchus (Lauprasert et al., 2009) . For all other OTUs for which this feature could be scored, the pattern of sculpting did not change between the main body of the squamosal and the immediate postorbital region.
(S8*) Ratio of forelimb to hindlimb length high (0.8) (C180.2): This feature is diagnostic for Alligatorellus beaumonti amongst all OTUs for which this character could be scored. Atoposaurus oberndorferi and Alligatorellus bavaricus both have similar ratios, 0.78 and 0.76, respectively, but Atoposaurus jourdani is distinct, with a ratio of 0.63 ( Table 3). However, this character state could not be scored for Theriosuchus , or the majority of our outgroup taxa, because of the relative rarity with which these specimens preserve associated and complete limb material. Therefore, although these unusual ratios are diagnostic amongst atoposaurids, we cannot determine whether they are unique or only local autapomorphies.
(S9) Ratio of tibia to femur length low (0.9) (C300.0): The relative dimensions of the tibia and femur are a feature that is closely shared with MB.R.3632 (0.91), Hoplosuchus (0.92), Alligatorium meyeri (0.93), and Atoposaurus jourdani (0.94). This ratio far exceeds that for ‘ Alligatorium ’ franconicum (0.64), and is distinct from Alligatorellus bavaricus (0.96) ( Table 3). This feature was also noted by Tennant & Mannion (2014), but those authors used a ratio of femur to tibia length.
(S10*) Nuchal osteoderms reduced, <50% of the size of the dorsal osteoderms (C307.1): This feature is distinct from the condition in Alligatorellus bavaricus , Alligatorium meyeri , Montsecosuchus , and Protosuchus , in which the preserved nuchal osteoderms retain the same size and morphology as the dorsal series, or only decrease slightly.
(S11*) Dorsal keel in dorsal osteoderms shifts laterally in more posterior dorsal osteoderms (C317.1): The position of the dorsal keel on the dorsal osteoderm series is distinct from that in Alligatorellus bavaricus , in which the morphology is more consistent along the axial column (Tennant & Mannion, 2014). This feature is not present in any other of the OTUs that we sampled, and therefore we consider it to be diagnostic for Alligatorellus beaumonti .
(S12*) Lateral ridge on sacral osteoderms forms an incipient posterior projection: The posterior development of the lateral keel (as noted in S9) into an incipient lateral projection amongst the more sacrally positioned osteoderms is diagnostic for Alligatorellus beaumonti . In Alligatorellus bavaricus , the morphology of the keel does not change anteroposteriorly (Tennant & Mannion, 2014), and Theriosuchus , Alligatorium meyeri , and higher neosuchians do not seem to possess this keel at all. ‘ Alligatorium ’ franconicum and Hoplosuchus are convergently similar, in that the lateral keel appears to form an anterolateral process, distinct from the ‘peg and socket’ articulation described for goniopholidids and T. guimarotae (Schwarz & Salisbury, 2005) . We did not incorporate this as a character to avoid duplication and overweighting of the observation that the morphology of the dorsal keel changes axially in Alligatorellus beaumonti (S11).
(S13*) Secondary osteoderms present in caudal series (C328.0): This feature does not appear to be present in any other atoposaurid that preserves caudal osteoderms.
Additional comments
In the holotype specimen of Alligatorellus beaumonti , the posterior-most maxillary teeth have a more labiolingually compressed, apically pointed morphology than the remaining teeth, similar to the ‘lanceolate’ morphology exhibited by several species of Theriosuchus (Schwarz & Salisbury, 2005; Lauprasert et al., 2011) and Brillanceausuchus , as well as the bernissartiid Koumpiodontosuchus (Sweetman et al., 2015) . This is different to the homodont dentition typically reported for Alligatorellus , which is usually described as possessing simple pseudocaniniform teeth that are smooth and lack ridges or carinae (e.g. Buscalioni & Sanz, 1990a,b; Thies et al., 1997). However, we do not assign this as a local autapomorphy of Alligatorellus beaumonti as it is only visible for one or two teeth in a specimen that shows strong evidence of dorsal compression. Its validity therefore requires further investigation pending the discovery of more specimens of Alligatorellus . Although this more lanceolate morphology was also figured for the posterior teeth of Alligatorellus bavaricus by Wellnhofer (1971), we have been unable to personally validate this on the figured specimen, and it is not visible on the holotype. Therefore, we urge caution in interpreting Alligatorellus as possessing lanceolate posterior teeth that are homologous to those found in Theriosuchus . Re-running our phylogenetic analysis [excluding ‘ Alligatorellus ’ sp. (MB.R.3632) and Pachycheilosuchus as before] with Alligatorellus scored as possessing lanceolate posterior teeth, we achieve a single MPT with a length of 793 steps with an unchanged topology. However, the presence of a lanceolate dentition instead becomes the basal condition in the clade containing atoposaurids and higher neosuchians, secondarily lost in Alligatorium meyeri , Brillanceausuchus , ‘ T. ’ ibericus , ‘ T. ’ sympiestodon , and the clade containing Koumpiodontosuchus and coelognathosuchians.
ALLIGATORELLUS BAVARICUS WELLNHOFER, 1971 (RE- RANKED BY TENNANT & MANNION, 2014) ALLIGATORELLUS BEAUMONTI BAVARICUS WELLNHOFER, 1971
Type locality and horizon
Solnhofen Formation, early Tithonian (Late Jurassic, Hybonoticeras hybonotum zone); Eichst att €, southeast Germany.
Type specimen
BSPG 1937 I 26 , a near-complete skeleton including the skull, lacking only the left forelimb, compressed onto a slab of lithographic limestone. Note that Tennant & Mannion (2014) incorrectly stated that the specimen number was LMU 1937 I 26.
Referred specimen
A specimen held in the private collection of E. Schopfel was described and referred to Alligatorellus bavaricus by Wellnhofer (1971), from the Wintershof Quarry (Solnhofen Formation, Eichst att €, southeast Germany).
Revised diagnosis and discussion
(S1) Concave profile of dorsal surface of snout in lateral view (C8.0): This feature represents a reversion back to the plesiomorphic state known for Protosuchus (Colbert & Mook, 1951) and Hoplosuchus (Gilmore, 1926) , with other atoposaurids and neosuchians usually presenting a straight profile in lateral aspect (with exceptions such as the longirostrine goniopholidid Amphicotylus ; Mook, 1942).
(S2) Small, slit-shaped antorbital fenestra, enclosed by nasals (C13.0 and C14.1): Alligatorellus bavaricus appears to possess a small, slit-like antorbital fenestra, similar to the notosuchians Gondwanasuchus (Marinho et al., 2013) and Malawisuchus ( Gomani, 1997) . In other taxa with an antorbital fenestra, including T. guimarotae (Schwarz & Salisbury, 2005) , T. pusillus , and ‘ T.’ ibericus , it is proportionally larger and rounded. Alligatorellus beaumonti does not appear to possess an antorbital fenestra, although part of the snout is embedded in the counterpart slab, with a small opening observable near the posterior margin of the nasals, which could be a diminutive fenestra. The presence of an antorbital fenestra is documented in basal crocodyliforms, including the protosuchians Hoplosuchus (Gilmore, 1926) and Protosuchus haughtoni (Gow, 2000) , Zosuchus (Pol & Norell, 2004a) , and thalattosuchians (Leardi
et al., 2012), but becomes closed in shartegosuchids, including Fruitachampsa (Clark, 2011). Pachycheilosuchus might have also possessed an antorbital fenestra (Rogers, 2003). (S3*) Extremely narrow and short skull, with a low skull width to orbit width ratio (<2.0) (C28.0): This character state is the lowest value for all atoposaurids, and much lower than all other OTUs in which this character was measurable. For other atoposaurids, this ratio is between 2.15 ( Alligatorellus beaumonti ) and 2.69 ( Atoposaurus jourdani ), and the only other taxon that comes close to this range is Wannchampsus (2.77). Brillanceausuchus , Montsecosuchus , and Theriosuchus species all have ratios between 3.0 and 3.5, with the ratio being considerably greater in longirostrine taxa and protosuchians ( Table 3). It is likely that this character state is influenced by ontogeny (Joffe, 1967), but the broad distribution of ratios amongst the sampled OTUs, which possess a range of body sizes and ontogenetic states, means that ontogeny is unlikely to entirely control this feature. (S4) Skull anteroposterior length to supratemporal fenestra length ratio ~7.2 (7.18) (C29.2): The proportional length of the external supratemporal fenestra is similar to Wannchampsus kirpachi (7.53), but distinct from Alligatorium meyeri (6.43) and Alligatorellus beaumonti (6.23), which have proportionally larger external supratemporal fenestrae. Consequently, we consider the proportionally short anteroposterior length of the supratemporal fenestra to skull length to be diagnostic for Alligatorellus bavaricus , because in Montsecosuchus this ratio is considerably higher (8.9) ( Table 3), with a much smaller supratemporal fenestra. (S5*) Posterior surface of nasals longitudinally crenulated (C69.0): The longitudinal crenulations on the dorsal surface of the nasals are not known in any other crocodyliform, in which the nasals are dorsally flat and sculpted like the rest of the cranial dorsal surface. (S6*) Smooth anterior region of parietal dorsal surface with a transverse frontal–parietal ridge, and shallow emargination at the posterior parietal–squamosal contact (C117.1) that develops into a thin dorsal groove connected to the supratemporal fenestra (C147.1): The morphology of the parietal is diagnostic, with a small anterior concavity at
the posterodorsal suture contact between the parietal and squamosal, leading to a shallow sulcus along this contact into the posterior margin of the supratemporal fenestra, and a smooth anterior dorsal surface. This is distinct from the condition observed in Theriosuchus in which this contact is deep and expands mediolaterally towards the supratemporal fenestra border, and from Alligatorellus beaumonti and Alligatorium meyeri in which the grooved contact is bordered by raised crests. The presence of a transverse ridge at the parietal – frontal suture distinguishes Alligatorellus bavaricus from all other species, in which this suture is flat.
(S7) Squamosal posterolateral lobe absent (C139.1):. The squamosal posterolateral lobe is completely absent in Alligatorellus bavaricus , a feature considered to be diagnostic amongst all atoposaurids.
(S8*) Distinct ridge on proximodorsal edge of scapula (C280.1): The scapula of Alligatorellus bavaricus can be distinguished from Alligatorellus beaumonti and other atoposaurids based on the presence of a distinct ridge on the proximodorsal surface. In all other specimens we analysed, the proximodorsal edge of the scapula is flat in lateral view, and confluent with the scapular shaft.
(S9) Extremely low radius proximodistal length to humerus length ratio (0.69) (C288.1): The radius to humerus ratio is extremely low, distinct from other atoposaurids in which the value is closer to 1.0. This low ratio is identical to that for ‘ Alligatorium ’ franconicum and Karatausuchus (Storrs & Efimov, 2000) , but higher than in Pachycheilosuchus (0.58).
(S10*) Low radius proximodistal length to tibia length ratio (0.64) (C289.1): This value is almost identical to that for Montsecosuchus , Pachycheilosuchus , Atoposaurus jourdani (0.61), and Protosuchus richardsoni (0.63), but much higher than that for Karatausuchus (0.47) and T. pusillus (0.55). Other taxa have proportionally long radii, including Alligatorium meyeri (0.74), Alligatorellus beaumonti (0.71), ‘ Alligatorium ’ franconicum (0.89), and Brillanceausuchus (0.88).
(S11*) Dorsal osteoderms with longitudinal medial ridge, becoming more laterally placed anteriorly (C311.1 and C312.1): This feature pertains to the morphology of the dorsal osteoderm series, which are distinct from those in Alligatorellus beaumonti (Tennant & Mannion, 2014) , as well as MB.R.3632 (Schwarz-Wings et al., 2011). In Alligatorium meyeri and T. pusillus , there is no evidence of a lateral keel.
| BSPG |
Bayerische Staatssammlung fuer Palaeontologie und Geologie |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |