
Microtyphlus charon, Ortuño & Sendra, 2011
|
publication ID |
https://doi.org/10.11646/zootaxa.2862.1.3 |
|
persistent identifier |
https://treatment.plazi.org/id/038387D9-1F6E-FFCA-FF3E-F882D17CFE32 |
|
treatment provided by |
Felipe |
|
scientific name |
Microtyphlus charon |
| status |
|
Description of Microtyphlus charon View in CoL n. sp.
(figs 1, 2a, 3d, 4d, 5e–f, 6a–b)
Holotype. 1 ♂, Spain, Valencia, Chera , “Surgencia de l’Orao” cave, [30SYJ155961, MGRS Datum: EUR-7]; 18- XI-2007; A. Sendra & S. Teruel leg. (VMO/ UA coll.).
Paratypes. 1 ♂, Spain, Valencia, Chera , “Surgencia de l’Orao” cave, [30SYJ155961, MGRS Datum: EUR-7]; 18-XI-2007; A. Sendra & S. Teruel leg. (VMO/ UA coll.) . 2 ♀♀, idem; 22-III-2005; A. Sendra leg. (VMO/ UA coll. & MVHN coll.) .
Etymology. The name of the specific epithet refers to the boatman of the Hades Kingdom (Charon, gr. Χάρων).
Diagnosis. The new species shows the general characteristics of the tribe Anillini . Blind (fig. 1) and apterous species. Depigmented integument, showing isodiametric micro-sculpture and a sparse covering of short setae on head, pronotum, and elytra. Pronotal setae inserted close to posterior angles. Elytra complete, elliptical, and convex, covering almost the entire abdomen (fig 1). Male first anterior tarsus slightly dilated. Body length (from mandible apex to elytron apex) 1.74–1.88 mm.
Description. Head (fig 1) moderately slender (ratio l/ w 0.97). Mandibles slender and prominent. Labial and maxillary palps typical of bembidiine beetles. Labium with complete labial prebasilar suture and six setae (two large setae near labial tooth) (fig 2a). Labrum trapezoidal. Antennae filiform (figs 1, 3d) with the half distal slightly moniliform; setulate and covered with tomentum from 2 nd to 11 th antennomeres. Ocular area smooth, lacking eyes and ocular scars. Cephalic disk with a scarce micro-pubescence, more sparse in comparison to pronotum and elytra. Cephalic chaetotaxy represented by one seta in the mandible scrobe, six labral setae (longer on each side), one seta at each side of clypeus, one seta on distal side of each front sulci, and two supraocular setae (anterior and posterior).
Pronotum cordiform (fig 1), slightly wider than longer (ratio w/l 1.25). Edges rounded in the two anterior thirds and sinuate in the posterior third. Anterior angles poorly defined, posterior ones slightly rounded and obtuse. Pronotal disk crossed by a shallow longitudinal sulcus. Lateral margin wide and regular. Pronotal chaetotaxy made up by two marginal setae, the anterior one slightly forward at the distal quarter, the posterior one near to the angle.
Elytra (fig 1) elliptical and convex, covering almost the entire abdomen. Ratio l/ w 1.6 (maximum width approximately at middle). Humeral region rounded, shoulders slightly prominent. Lateral margin slightly serrate on the two anterior thirds and with short setae. Scutellum large and triangular, without developed striae. Elytron chaetotaxy made up by a parascutellar pore located at level with basal margin, three discal setae (anterior at level with the hiatus between the 2 nd and 3 rd umbilicate setae; middle seta at level with the 4 th umbilicate seta; posterior seta ahead of the 7 th umbilicate seta), umbilicate series made up of nine setae. Second, 6 th, and 9 th setae very long. First three setae gathered together and equidistant, 4 th seta very distant from the 3 rd and from 5 th and 6 th pairs (which are very close); 7 th, 8 th, and 9 th setae making up a triangle (without geminate pair), apical seta almost aligned with posterior disk seta.
Legs (fig 1) longer than average in other Iberian Anillini genera. First protarsomere slightly dilated and ventrally covered by adhesive phaneres in the male, no other secondary sexual dimorphism on legs. Mesal surface of protibia with a cleaning organ. Both meso- and metatarsi strongly setulose and with a markedly elongated first tarsomere (fig 4d), slightly longer that the sum of the distal four.
Abdomen ventrally setulose. Last ventrite of female with four long setae near apical margin, in male only two.
Aedeagus (figs 5e–f) with a short, voluminous median lobe; most sclerotized areas are found in the internal piece, the basal lamina, the apex, and the region where the parameres are articulated. Basal lamina bent towards clearly raised apex. Apex long and turned to right in dorsal view. Median lobe poorly sclerotized, particularly at base. Internal sac voluminous, mainly membranous, with a long helminthoid-shape sclerite and a dorsal sulcus in anterior position. Parameres asymmetrical, the right one smaller and both bisetulose.
Genital shield of female genitalia (fig 6a) are made up of three pieces at both sides of the plane of symmetry: dimerous IX-gonopods (gonocoxite + gonosubcoxite) plus an IX-laterotergite. IX-gonocoxite unguiform and heavily sclerotized, with two thick and long setae inserted close to external edge, one located on ventral surface, the other on dorsal surface. Mid gonocoxite showing a foseta close to the internal edge, in which a thin, long sensory seta is housed. IX-gonosubcoxite poorly sclerotized and lacking setae, almost 2 times longer than wide. IX-laterotergite wing-form, oblique, and slightly sclerotized, showing a membranous basal margin provided with fifteen medium-length spine-shaped setae. Spermathecal complex partially membranous (fig 6b). Spermatheca duct short (0.3 mm), sinuous, and wide in diameter all along its length. Spermatheca perceptibly sclerotized, becoming bacilliform, almost rectangular (ratio l/w 2). Spermathecal gland large and sclerotized in distal half and partially on proximal half, opening into middle region of seminal receptacle.
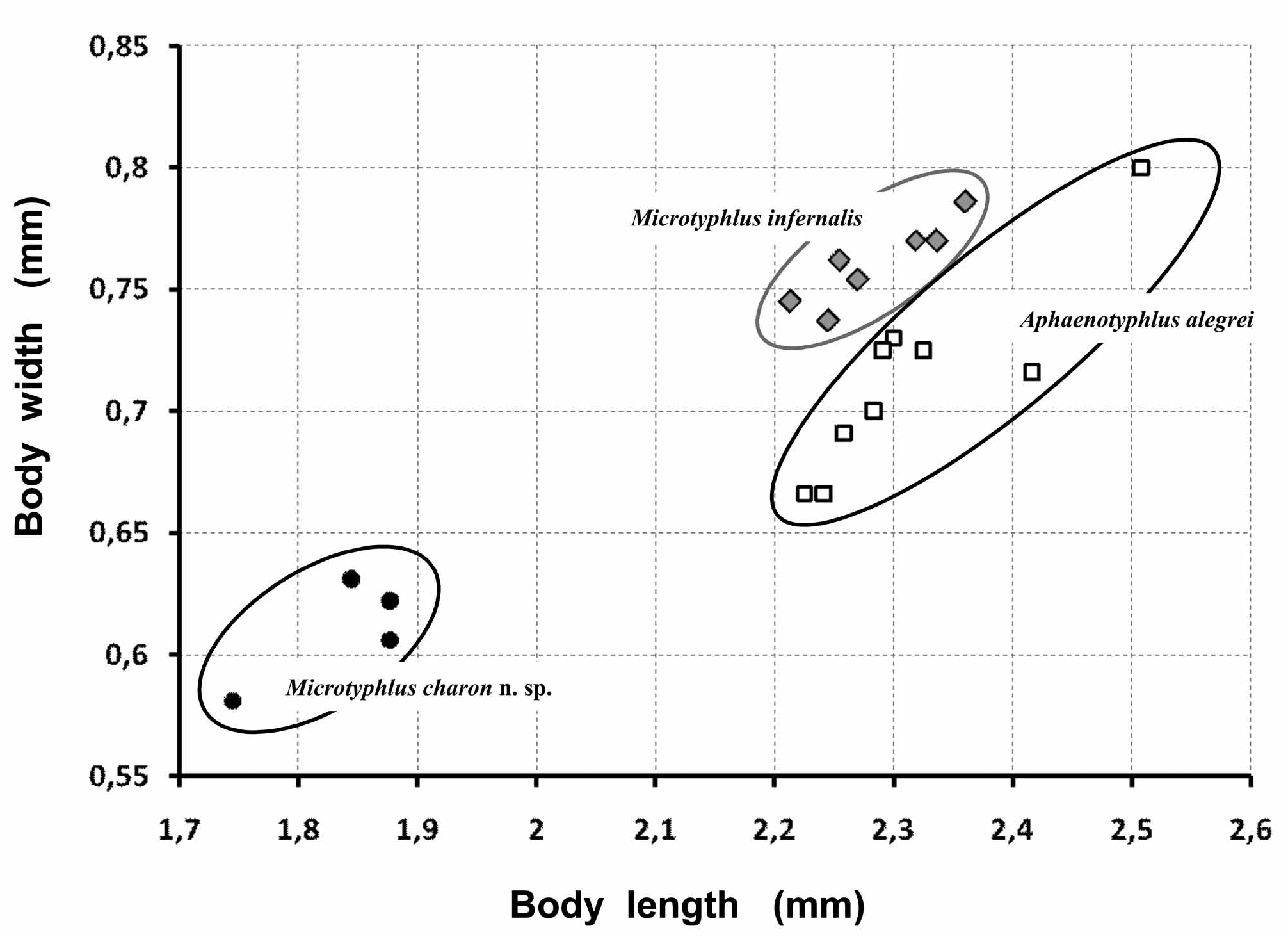
Affinities: M. charon n. sp. is close to M. infernalis Ortuño & Sendra 2010 according to general body shape (proportions and relative size of head, pronotum, and elytra), the remarkable convexity, the chaerotaxy (setae number and topology), and hypogean life. At first sight the most conspicuous difference between both species is the smaller body size of M. charon n. sp. ( Fig. 7 View FIGURE 7 ). Average body length is 2.285 mm in M. infernalis , whereas the holotype of M. charon n. sp. is 1.877 mm. Similar results are found in body width ( Fig. 7 View FIGURE 7 ). More subtle differences are found in the shape of the posterior angles of pronotum, which are straight and sharp in M. infernalis and obtuse and rounded in M. charon n. sp. Both species share the same model of aedeagus but show differences in the development of the apical lamina and the degree of sclerotization of the internal sac (figs. 5c–f).
Body size is even larger and wider in Aphaenotyphlus alegrei (n= 9; average length 2.316 mm; fig. 7).
Habitat: M. charon n. sp. has been only found in the type locality, the “Surgencia de l’Orao” Cave. This cave is located on the southwestern slope of the mountain “Cumbre del Burgal,” Chera district, in the province of Valencia (eastern Spain). This natural cave is formed of Lower Cretaceous (Aptiense) limestone with thin clay layers that act as an impermeable baseline for this small, short cave, less than fifty metres long in total (fig 8). This cave has a natural lower entrance, where water flows out during flood periods, and an artificial entrance, dug out to make water extraction easy in dry periods. The water is extracted by an electric pump powered by a solar panel and pumped out from a small siphon at the end of the main gallery, which is flooded all year round. The main gallery remains flooded during weeks or even months in rainy periods with the exception of the upper zone, where the ceilings of the gallery are higher, at about 25 m from the natural entrance (fig 8). We could see most of the hypogean species in that section of this particular cave and it is where the four individuals of M. charon n. sp. were collected, always under stones. Temperature of the room section oscillated between 13.6–14.6 ºC.
M. charon n. sp. lives in a community comprising a number of detritivores such as Cordioniscus stebbingi (Patience 1907) ( Isopoda : Styloniscidae ); a troglophilic species in dry areas and four troglobitic species abundant, one species of Collembola of the Entomobryidae family, one species of Coletinia sp. ( Zygentoma : Nicoletiidae ), Campodea aff. agena Condé 1951 ( Diplura : Campodeidae ), and a leiodid beetle Anillochlamys sendrai Fresneda & Salgado 2006 ( Coleoptera : Leiodidae ).
| UA |
University of Alabama |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.