
Metallactus albipes Suffrian, 1866
|
publication ID |
https://doi.org/10.11646/zootaxa.4413.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:D65DAEB6-0499-44F9-A70D-0720BA71D520 |
|
persistent identifier |
https://treatment.plazi.org/id/03DB8796-3206-9800-FF48-1190FB1FFCE1 |
|
treatment provided by |
Plazi |
|
scientific name |
Metallactus albipes Suffrian, 1866 |
| status |
|
Metallactus albipes Suffrian, 1866
( Figs 1 View FIGURES 1–2 ; 4 View FIGURES 3–4 ; 22 View FIGURES 20–28 )
Metallactus albipes Suffrian, 1866: 366 ; Clavareau, 1913: 92; Blackwelder, 1946: 640. Metallactus nigrofasciatus Suffrian, 1866: 363 ; Fiebrig, 1910: 248; Clavareau, 1913: 93; Blackwelder, 1946: 641; White, 1975: 88; Riding, 1983: 127. n. syn.
Type material. Metallactus albipes : in MNHUB there are 20 specimens ( 15 ♀♀ and 5 ♂) labeled: “ Syntypus Metallactus albipes Suffrian, 1866 labelled by MNHUB 2013” [Red label, printed]. Among them, only a few specimens carry an ancient blue handwritten label which reports the locality in abbreviated form: “Allegr. Sellow”; “Cassapava Sell.”; “Porto all Brasil Sellow”; “ Brasil Sellow”; “ Brasil Virm.”; “Cassap. Sellow”. All the other specimens carry only a green printed recent label: // “Hist.-Coll ( Coleoptera ) Nr. 23987 Metallactus albipes Suffrian var. Brasil., Sello, Virmnd. Zool. Mus. Berlin” //. The number of specimens and the label information match the registration data from the old catalogue of the MNHUB (“23987 albipes Suffr var 20 exx. Brasil, Sello, Virmd”). Therefore, there is no doubt that this is the complete Berlin’s type series studied by Suffrian. The examination of the aedeagus revealed that one out of the males actually belongs to a different species ( M. verrinus Suffrian, 1866 ). Even among females there is a considerable variability and it is possible that some of them do not belong to the same species. Besides, following Suffrian's statements and on the basis of the label data, 9 putative syntypes ( 1♀ coll Baly, 8♀♀ coll Clark) have also been detected in BMNH. As a matter of fact, only the specimen from Baly collection has been proved to fit in well with all the data of the original description (“ Metallactus albipes Suffr Brasil Type Suffr Coll Deyrolle” [white label, handwritten] // “Type Suffr Coll Deyrolle” [white label, handwritten] // “Baly coll.”[white label, printed] // “ Syntype ”[white with blue border, printed]) and therefore it has been designated as paralectotype. The typification has been made as follows. LECTOTYPE: ♂, aedeagus extracted and glued, together with the abdomen, on the same label bearing the specimen, // “Allegr. Sellow” [blue label, handwritten] // “ Syntypus Metallactus albipes Suffrian, 1866 labelled by MNHUB 2013” [red label, printed] // “IDe_176” [white label, handwritten] // “albipes 161 syntype 1 endof” [red label, handwritten] // “ Metallactus albipes Suffrian, 1866 LECTOTYPUS D. Sassi des.” [red label, printed] // ( MNHUB). The remaining syntypes have been labelled as PARALECTOTYPES (// “ Metallactus albipes Suffrian, 1866 PARALECTOTYPUS D. Sassi des.” [red label, printed] // ( MNHUB and BMNH).
Metallactus nigrofasciatus: Suffrian claimed that there were no male specimens in the type series. I could track down three ♀♀ in BMNH and two ♀♀ in MLUH which fit well with the data in the original description. The typification has been made as follows. LECTOTYPE: ♀, pinned, // “ Type Suffrian coll Baly ” [white label, handwritten] // “ Baly coll.” [white label, printed] // “ Metallactus nigrofasciatus Suffr Brazil ” [white label, handwritten] // “ Syntype ” [white with blue border, printed] // “ Metallactus nigrofasciatus Suffrian, 1866 LECTOTYPUS D. Sassi des.” [red label, printed] // ( BMNH). PARALECTOTYPES: 1♀, same data of the lectotype ( BMNH); 1♀, “a D. Suffrian determinat. ” [white label, printed] // “67.56” [white label, printed] // “ Brazil ” [white label, printed] // “ Syntype ” [white with blue border, printed] ( BMNH); 2♀♀, pinned, “24718” and “20419” [white label, handwritten] // “nigrofasciatus m. Brasil” [blue label, handwritten] // “MLU Halle WB Zoologie S.Nr. 71116” [white label, partly printed]. All paralectotypes were labelled “ Metallactus nigrofasciatus Suffrian, 1866 PARALECTOTYPUS D. Sassi des.”[red label, printed] // ( BMNH and MLUH) .
Type locality. Metallactus albipes: Porto Alegre ( Rio Grande do Sul, Brazil). Metallactus nigrofasciatus : Brazil.
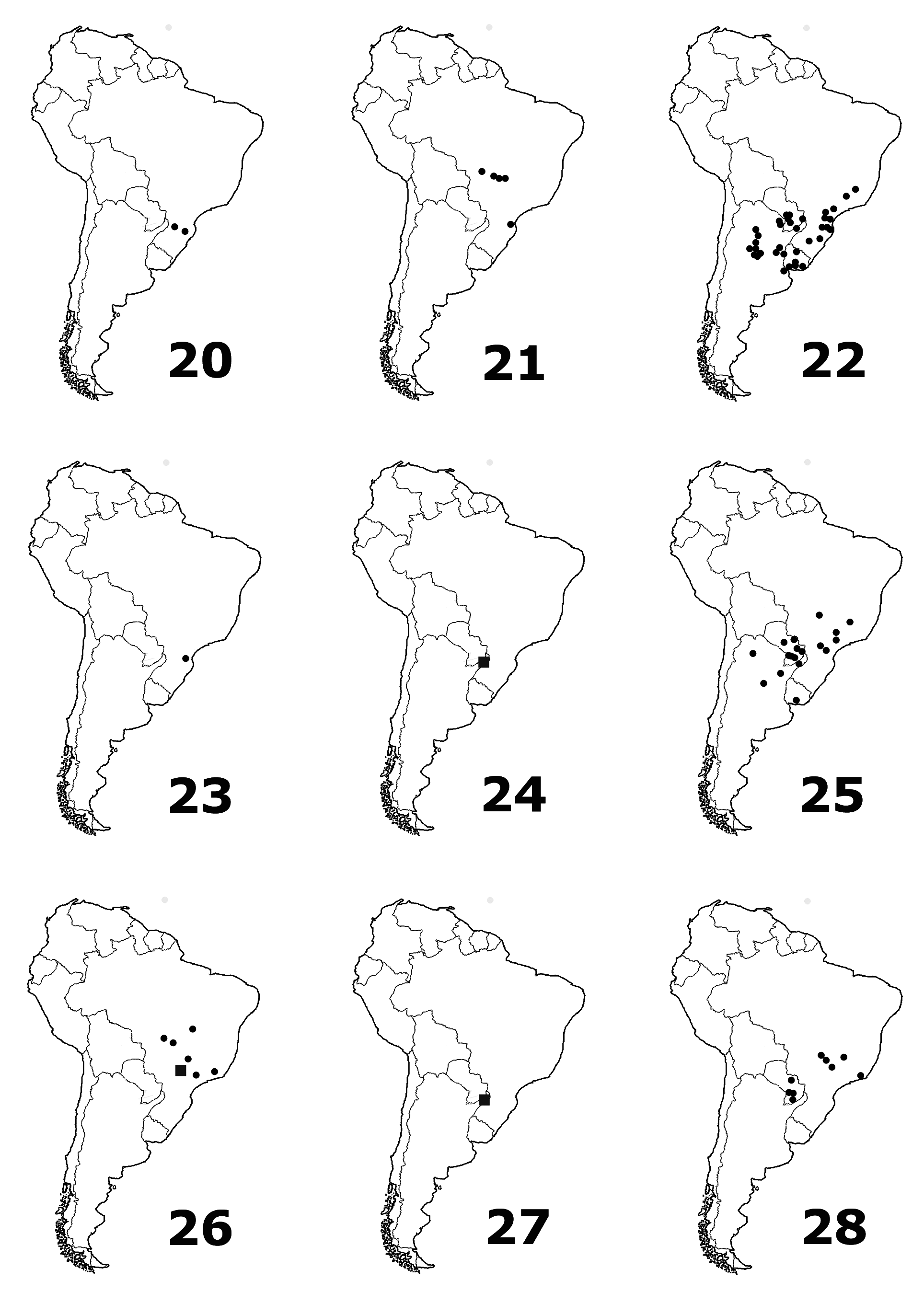
Further material examined. BRAZIL: Vacaria , Rio Grande do Sul 12.I.1974 McFayden & La Roca (5, USNMNH) ; Puerto Alegre Santa Cruz Rio Grande do Sul Hensel (3, MNHUB) ; Rio Grande do Sul Hamburgo Velho Ertl (1, ZSM) ; Campos Gerais (2, MHNH) ; Caraça Minas Geraes E. Gounelle 12.1885 (1, MHNH) ; São Paulo (1, NHMP) ; São Paulo Cantareira 17.XI.1929 Halik (1, USNMNH) ; Florianopolis 28.III.1963 on Baccharis spicata F. D. Bennett (2, BMNH) ; Parana Curitiba XII.1940 J. Guerin (1, USNMNH) ; Santa Catarina Laguna 27.II.1979 (1, BMNH) ; Santa Catarina 20 mi. E Campos Novos 1.III.1980 D. B. Thomas (1, ERPC) ; S. Catharina coll. H. Clavareau (1, MHNH); Palmeira Paranà D. F. Grillo (1, MSNG) ; Brazil ex coll. Chevrolat gift F.C. Bowditch (1, USNMNH) ; ” Brasilia ” (1, MNHUB) ; “ Brésil ” Bryol (1, NHMP) ; “ Brasilien ” (1, ZSM) . PARAGUAY: Alto Parana 45 Km W P. to Pres. Stroessner 28.I.1983 E. G. Riley (1, ERPC) ; Pres. Hayes 42 Km NW Benjamín Aceval 31.I.1983 E.G. Riley (6, ERPC, MSNG) ; Pres. Hayes 2 Km SW Villa Hayes 30.I.1983 and 6.II.1983 E. G. Riley (3, ERPC) ; Asuncion (2, NHMP and NMV) ; Asuncion 27.X.1904 Vezényi (4, HNHMB) ; Asuncion Vezényi (2, HNHMB) ; Asuncion (1, USNMNH) ; Asuncion Revoil 1891 (8, MHNH) ; Sapucay W. T. Foster (3, USNMNH) ; San Bernardino W. Eisenlehr (2, MNHUB) ; San Bernardino Fiebrig (2, NMV) ; Hohenau III.1953 Walz (1, USNMNH) ; Paraguay Central P. Germain 1885 (1, MHNH) ; Paraguay C. Fiebrig (3, MNHUB) ; Paraguay (1, HNHMB) ; Paraguay (1, USNMNH) . ARGENTINA: Cordoba R.A. Huerto Grande 1000m X 1952 (4, NHMB) ; Cordoba Dep. Calamuchita El Sauce XII 1938 Manuel Viana (5, USNMNH) ; Cordoba Cuadras 29.I.1953 (6, USNMNH) ; Cordoba La Falda I.1945 coll. Monrós (1, USNMNH) ; Cordoba La Falda X.1974 Viana (3, DSPC) ; Cordoba Casabamba I.1946 coll. Monrós (1, USNMNH) ; Cordoba Capilla del Monte XII.1945 coll. Monrós (1, USNMNH) ; Cordoba Capilla del Monte Hosseus (1, ZSM) ; Cordoba Fanti Sierra de Cordoba 11.I.1966 Mahunka (1, NHMP) ; Cordoba Anizacate Maristas (1, USNMNH) ; Cordoba La Plata (1, USNMNH) ; Cordoba Sierra Grande 1000–1200m Los Hornillos 12–14.XI.1999 Zabransky (1, DSPC) ; Sierra Cordoba 5.XI.1927 (1, NHMP) ; Territ. de Misiones (2, NHMP) ; Territ. de Misiones Le Moult Coll. Wagner (3, NHMP) ; Misiones Haut-Parana San Ignacio (1, MNHUB) ; Misiones Haut-Parana San Ignacio Le Moult Coll. Wagner (3, NHMP) ; Territ. de Misiones Rio Parana Le Moult Coll. Wagner (3, NHMP) ; Misiones Rio Parana (3, NHMP, HNHMB) ; Misiones Pastoreo Grande II.1954 Walz (1, USNMNH) ; Territ. de Misiones Rio Parana (1, MNHUB) ; “des Missions ” (1, MHNH) ; Entre Rios Liebig XII.1987 (1, DSPC) ; Entre Rios Paranà General Racedo 8.I.1954 Fritz (1, USNMNH) ; Umg . Buenos Aires 13.I.1931 (2, NHMP) ; Formosa Gran Guardia XII.1951 Föerster (1, USNMNH) ; Formosa 34 Km NE Formosa 27.I.1989 C. W. & L. W. O'Brien & G. J. Wibmer (1, BYU) ; Santa Fé Garay (4, USNMNH) ; Chaco de Santiago del Estero Env. D'Icano Mistol Paso E.R.Wagner 1911 (1, MHNH) ; Santiago del Estero Ojo de Agua III.1992 (1, DSPC) . URUGUAY: Montevideo So. Am. Paras. Lab. No. 877.4 x.1942 on Baccharis spicata, Parker Berry Silveira (5, USNMNH) ; Rivera Ruta 30 Subida de Pena 9.V.1968 Günter & Carbonell (3, MNHUB) ; Rocha 65 Km SW Rocha 7.II.1989 C. & L. O'Brien & G. Wibmer (1, DSPC) ; Colonia San Pedro Ruta 21 ca. Km 186.5 10-II-2014 G. J. Wibmer (1, DSPC) ; Canelones, 19 Km NE Canelones R 63 7.II.1989 C. & L. O'Brien & G. Wibmer (1, BYU) ;
Distribution. Argentina, Brazil, Paraguay, Uruguay. New to Paraguay and Uruguay.
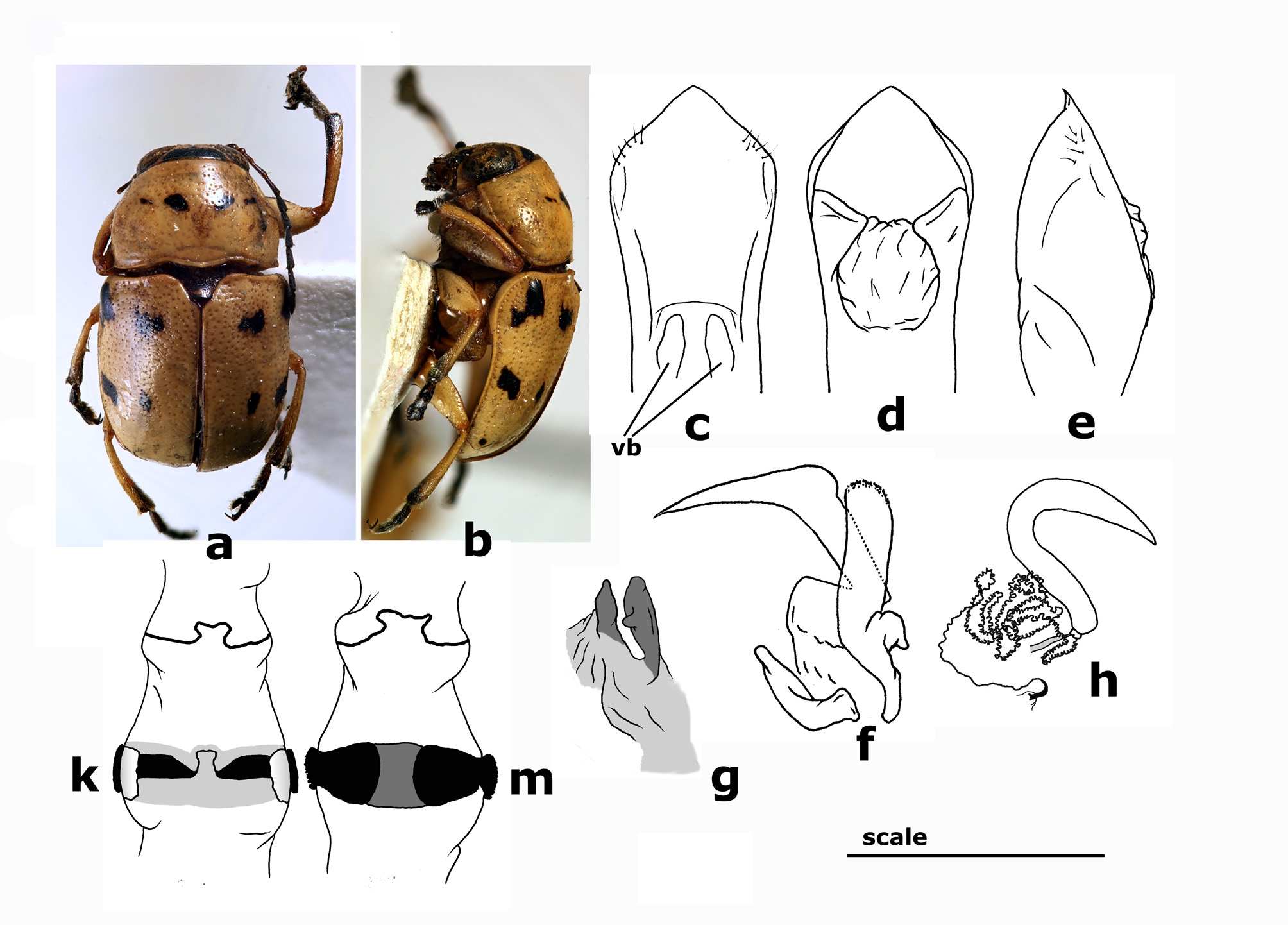
Diagnosis. A Metallactus of large size, especially variable in dorsal chromatic pattern. The most distinctive trait is the shape of the branches of sclerite IV of endophallus, which are concave at apex (= saddled) ( Fig. 4f View FIGURES 3–4 ). This morphology has been observed only in specimens with general habitus consistent with this species, and has been considered here the key factor for the attribution to M. albipes , regardless of the chromatic appearance. Aedeagal apex is sometimes delimited at base by a pair of barely perceptible lateral thickenings ( Fig. 4c View FIGURES 3–4 ), this trait is not always perceivable, but seems to be apparent in this species and lacking in other ones of similar habitus, being of some help in diagnostic tasks. M. albipes most resembles M. corruptus in habitus and dorsal coloration, but the latter species distinctly differs in smaller size, constant lacking of lateral thickenings at base of aedeagal apex and different shape of sclerite IV of endophallus. M. guarani is also similar but it differs in smaller size, slimmer body, lighter dorsal coloration, abdominal ventrites completely yellow and for the presence of two weak oblique bulges close to the aedeagal basal foramen ( Fig. 19c View FIGURE 19 ). Dorsal spicule ( Fig. 4g View FIGURES 3–4 ) seems to be quite variable in shape, sometimes being strongly reduced, or apparent, conical and fairly pigmented. However, giving the present state of the art it has been impossible to detect any putative taxonomic significance linked to this variability.
Description of male. Habitus in Fig. 4 View FIGURES 3–4 a–b (LT). BL = 5.5–6.2 mm, BW = 3.4–3.8 mm, PL = 1.9–2.1 mm, PW = 3.2–3.4 mm. Interocular distance 11.5–12.5% of BL.
Head yellow with vertex, upper part of frons, canthus of eyes and insertion of antennae black. Surface barely setose with few moderately impressed scattered punctures. Frons with a fairly impressed mid-cranial suture. Labrum yellowish or blackish with lighter outline. First five antennomeres yellowish, the subsequent darkened, rather dull and more diffusedly setose.
Thorax yellow with a transverse black median stripe not reaching the lateral margins. Median stripe can be reduced, split in two or four separate spots to almost completely obliterated. Transverse, trapezoidal, with narrow lateral margins almost regularly convergent towards anterior region. Posterolateral impressions barely perceivable. Surface moderately shiny, covered by scattered moderately impressed punctation.
Scutellum black, dull, distinctly raised with apex truncated or slightly rounded. Surface mostly glabrous, with tiny scattered punctures.
Elytra yellow with six black spots arranged in three tranverse pairs. First pair of spots close to basal margin. Second just behind the middle. Third one, smaller, on the apical clivus. Inner spot of second pair generally more transverse than the other ones. Spots can be laterally expanded to join to each other into transverse bands (form “ nigrofasciatus ”, Fig. 4a View FIGURES 3–4 1 View FIGURES 1–2 ), but they can also be fairly reduced or missing and in some cases elytra are completely yellow. Parallel-sided, base and middle equally wide, regularly convex, inner side of humeral calli weakly impressed. Post-scutellar area very weakly raised. Surface moderately shiny with small and shallow punctures arranged in very irregular rows, disappearing on elytral apex. Intervals smooth.
Surface of pygidium yellow, smooth, haired, shallowly and sparsely punctured.
Inferior parts blackish with lateral sides of first abdominal sternites, last abdominal sternite yellow. Surface coarsely punctured with scattered yellowish setae. Prosternal process moderately broad, surface coarsely punctate, covered by long and whitish setae, longitudinally sulcate, apex rounded or subtriangular, blunt, devoid of apical denticle. Legs yellow with tarsi, external side of anterior femora and apical half of tibiae more or less darkened.
Fifth abdominal ventrite marked by a very feeble median depression, posterior margin weakly notched. Median lobe of aedeagus ( Figs 4 View FIGURES 3–4 c–e) cylindrical, rather stout, with apex short, in ventral view apex sometimes delimited by a pair of barely perceptible lateral thickenings. Ventral surface scarcely swollen, smooth, even, only few setae marking small hairy dents close to apex. In lateral view apex fairly bent ventrally.
Endophallus ( Figs 4 View FIGURES 3–4 f–g) with sclerite I weakly sclerotized with quite small but perceptible denticle. Dorsal spicule in many specimens reduced to a thin ribbon-like thickening in front of sclerite I, in other specimens spicule well developed, erect, small, conical in section. Sclerite II strongly developed, gradually tapering toward apex. Sclerite III short, gradually tapered towards apex, sometimes upper rim notched in the middle. Branches of sclerite IV subequal to or slightly higher than sclerite III in the folded up structure, concave at apex (= saddled), surface with developed micro-sculpture, consisting of a series of microdenticles along the apical margin, most evident in the ventral apical corner, and tiny oblique wrinkles developed along the outer surface of each branch.
Female. Habitus in Fig. 4a View FIGURES 3–4 1 View FIGURES 1–2 (PLT of M. nigrofasciatus ). BL = 6.3–7.8 mm, BW = 3.7–4.8 mm, PL = 1.9–2.4 mm, PW = 3.3–4.2 mm. Interocular distance 13.5–14.1% of BL.
Fifth abdominal ventrite in females with a rather deep hemispherical pit. Vasculum of spermatheca ( Fig. 4h View FIGURES 3–4 ) slender, elongated. Apex long and acuminate, often slightly curved downwards, sometimes straight or curved upwards, base vaguely curved backwards. Duct rather short, of uniform size, slender and coiled, coils rather close, gradually reduced in the last portion, which is almost straight; duct insertion on bursa copulatrix button-like or slightly lengthened, clearly pigmented. Rectal apparatus ( Figs 4 View FIGURES 3–4 k–m) with dorsal sclerites laterally not projected beyond rectus, narrow, tapered towards median line. Apodemes hyaline, bent upwards and leaning against rectum, thus barely visible from above. Ventral sclerite large, ribbon-like, slightly tapered on sides, less pigmented in middle, with rounded, crenulate apodemes wider than rectum.
Available biological data. Some information about the biology of the species (as M. nigrofasciatus ) was supplied by White (1975) and Riding (1983). The latter author reported the species to feed on Baccharis spicata (Lam.) Baill. and B. lymanii G.M. Barroso ex G. Heiden in Brazil. Some label data of examined specimens coming from Uruguay ( Montevideo) and from Brazil (Florianopolis) confirm Baccharis spicata as host plant.
Remarks. Suffrian (1866) considered this species particularly critical, and pieced together a rather heterogeneous type series regarding general habitus and colour pattern, that suggest it had been thought more like a “waste basket” assemblage rather than a clearly defined species. Besides, he described Metallactus nigrofasciatus ( Fig. 4a View FIGURES 3–4 1 View FIGURES 1–2 ) on females only and on the basis of chromatic differences consisting in a more developed black pattern on dorsal parts. Despite the abundant material examined, no constant peculiarities have been detected in elytral and pronotal color and punctuation allowing any congruent subgroup to be separated, all of these characters showing great variability. Not even the aedeagus morphology and female sexual characters appear to be of great help. Nevertheless, the shape of saddle-like apex of sclerite IV is a unifying feature that allows us to consider at a congruent specific level a group that would otherwise be arbitrarily treated on the basis of uncertain characters. The conformation of the apex of the sclerite IV, although showing some slight variation, is very characteristic and remains constant in a large number of the examined specimens, both in individuals chromatically assignable to the “ albipes ” form, and in those characterized by a “ nigrofasciatus ” design, and not showing any geographical significance. Considering the high colour variability, not correlated with any morphological character and the unifying factor given by the peculiar configuration of sclerite IV, it is considered appropriate to synonymize M. nigrofasciatus with M. albipes . The names M. nigrofasciatus and M. albipes were published together ( Suffrian 1866) and the latter is chosen as valid name of the species because of the presence of specimens of both sexes in the type series (art. 24.2 ICZN).
It should be noted that the size and shape of the dorsal spicule, which in other cases have been treated as clues of specific level, have not been considered relevant herein. The reason is that in this case the amount and the kind of data collected has not allowed me to operate differently. In fact, in most of the specimens the dorsal spicule is reduced to thin, ribbon-like and poorly sclerotized structures, but in others it is much more developed, conical in section and with an erect and well-defined apex. It does not seem that these different traits correlate with other relevant aspects, appearing both in darker specimens (“ nigrofasciatus ” form) and in completely yellow individuals (“ albipes ” form).
Finally, in a specimen coming from an ancient collection of the Museum of Munich, bringing the generic indication of origin “ Brasilien ” and having the dorsal parts yellow, with few black markings consisting of scutellum contour, suture, a small humeral and two apical rounded spots on elytra, the sclerite IV apex is hammerlike and not saddled. That means it is expanded, with the upper edge not concave but straight, and the anterior and rear angles fairly squared and not roughly pointed. Additionally, the dorsal spicule is short, conical and distinctly outlined. Besides, some external traits are quite unusual compared to what is observed in this species: black colour is completely missing on forehead and vertex; scutellum is predominantly yellow, regularly and finely edged in black; pronotum is less transverse and less bulging along the back edge and front sides of the pronotum are not converging; pronotal punctation is fairly robust, dense and slightly irregular ( i.e. of different sizes), especially on the center of the disc. The same hammer-like conformation of sclerite IV has also been observed in one male from Lagoa Santa, but in this case the chromatic drawing matches the most common M. albipes color pattern. Therefore, it can not be excluded that these two specimens belong to different species, but in the absence of additional data, it was preferred to consider them as outliers in a range of possible variation within the species.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Cryptocephalinae |
|
Genus |
Metallactus albipes Suffrian, 1866
| Sassi, Davide 2018 |
Metallactus albipes
| Riding, N. 1983: 127 |
| White, G. 1975: 88 |
| Blackwelder, R. E. 1946: 640 |
| Blackwelder, R. E. 1946: 641 |
| Clavareau, C. H. 1913: 92 |
| Clavareau, C. H. 1913: 93 |
| Fiebrig, K. 1910: 248 |
| Suffrian, E. 1866: 366 |
| Suffrian, E. 1866: 363 |