
Mesabolivar, GONZALEZ-SPONGA, 1998
|
publication ID |
https://doi.org/ 10.1206/0003-0090(2000)254<0001:NWPSAP>2.0.CO;2 |
|
persistent identifier |
https://treatment.plazi.org/id/03ACD276-8FE8-FF28-FF3E-FC7743E83C5C |
|
treatment provided by |
Felipe |
|
scientific name |
Mesabolivar |
| status |
|
MESABOLIVAR GONZÁLEZ-SPONGA, 1998 View in CoL View at ENA
Mesabolivari González-Sponga, 1998: 27 (type species by original designation M. pseudoblechroscelis González-Sponga, 1998 ; not examined—the newly described M. huambisa is a very close relative).
NOTES: This genus roughly corresponds to Blechroscelis Simon, 1893 . However, the type species of Blechroscelis ( Pholcus annulipes Keyserling ) is congeneric with the type species of Priscula Simon, 1893 , a genus described in the same paper as Blechroscelis . The first question that has thus to be answered is the following: is B. annulipes a misidentified type species? If yes, Article 70.3 of the International Code of Zoological Nomenclature (ICZN, 1999) would apply. I believe it is not, for two reasons: first, there is apparently no material labeled ‘‘ B. annulipes ’’ in the MNHN in Paris, meaning that Simon probably never saw any material of what he considered to be B. annulipes , but based his decision on Keyserling’s description; second, Keyserling’s (1877) description is accurate and detailed, so it is not probable that Simon missed the fact that B. annulipes did not quite fit his own description of Blechroscelis .
This means that Priscula and Blechroscelis have to be treated as subjective synonyms. Since they were proposed in the same publication, the Principle of the First Reviser (ICZN, 1999: Art. 24.2) provides that the first reviser determines the relative precedence. I chose Priscula as senior synonym, for two reasons: first, it leaves the well-defined genus Priscula intact in its original meaning; second, it only affects Blechroscelis , a polyphyletic genus that has come to include species from what are herein considered to be representatives of five genera.
The next question is: which is the next available name for the species corresponding to Simon’s original conception of Blechroscelis ? González-Sponga (1998) proposed the new genus Mesabolivari , whose type species clearly falls into the large group of species corresponding to Simon’s Blechroscelis . However, Mesabolivari has the form of a Latin two-word phrase (‘‘tableland of Bolívar ’’—I am indebted to H. D. Cameron for advice concerning this and what follows). The ICZN (1999: Art. 26) specifies that such a spelling must be considered Latin, and thus Mesabolivari is a two word-phrase, not a compound noun. It does not, therefore, fulfill one of the Criteria of Availability (ICZN, 1999: Art. 11.8), and must be corrected. [It is also not an ‘‘arbitrary combination of letters’’ (Art. 11.3), since it is explicitly meant to refer to the city Mesa Bolívar]. Instead of changing ‘‘bolivari’’ to the corresponding nominative singular (which would result in the grammatically awkward combination ‘‘Mesabolivarus’’), I here choose the emend- ed form Mesabolivar , which renders it a simple noun without the complication of Latin grammar.
The final question regards the gender of the genus. No gender was explicitly attribut- ed to Mesabolivari by González-Sponga (1998), and no gender is grammatically selfevident, neither in Mesabolivari nor in Mesabolivar . The specific name of the single species included by the author ( pseudoblechroscelis ) is a noun in apposition (ICZN, 1999: Art. 11.9.1.2) (anything ending in ‘‘-scelis’’ is a noun, not an adjective), and gives therefore no indication of the intended gen- der of the genus Mesabolivari . Therefore, Mesabolivar is to be treated as masculine (ICZN, 1999: Art. 30.2.4).
DIAGNOSIS: (The diagnosis given herein is valid only for a core-group of species close to the type species; see Specific Relationships below for species assigned tentatively). Medium-sized to fairly large (total length ~ 3–6 mm), usually dark, eight-eyed pholcids with long legs, usually oval to elongate opisthosoma; distinguished from similar neotropical genera ( Coryssocnemis , Carapoia ) by the median groove or pocket on the female epigynum, and a corresponding pair of contiguous apophyses frontally on the male chelicerae. These characters are missing in a few species ( M. huanuco , n. sp., togatus (Keyserling) , maxacali , n. sp.) which are nevertheless assigned to the core-group because of overall and specific similarities (see below).
DESCRIPTION: Total length usually ~ 3–6 mm. Carapace with distinct thoracic groove, ocular area moderately to conspicuously elevated, with eight eyes, AME smallest. Distance PME-ALE relatively large (65–100% of PME diameter). Sternum without anterior humps. Male clypeus unmodified. Basal segment of male chelicerae usually with pair of contiguous apophyses frontally, sometimes with additional apophyses more proximally; without modified hairs; fangs unmodified; without stridulatory ridges laterally. Male palpal coxa with retrolateral apophysis, procursus and bulb relatively simple, variable in shape. Tarsal organ exposed [examined: M. huanuco , n. sp., eberhardi , n. sp., aurantiacus (Mello-Leitão) : fig. 81, junin , n. sp., iguazu , n. sp., cyaneotaeniatus (Keyserling) ]. Legs long (leg 1 usually ~ 10–20 × body length; tibia 1 l/d usually ~ 70–100; 45 in M. botocudo , n. sp., 110 in M. aurantiacus ); leg 1 longest, leg 2 usually longer than leg 4, leg 3 shortest; in most species femora of some walking legs (usually femora 2, 2 and 3, or 3) thicker than others (not in M. huanuco , locono , n. sp., iguazu ); legs sometimes with spines on metatarsi, only in M. spinulosus (Mello-Leitão) and M. cambridgei (Mello-Leitão) with spines on femora and tibiae; without curved hairs (except in some M. cambridgei specimens), without vertical hairs (except M. cyaneotaeniatus , which has vertical hairs on femora); retrolateral trichobothrium of tibia 1 very proximal (at 1.5– 3%); tarsus 1 usually with 30 or more pseudosegments (~ 25 in M. iguazu and botocudo ). Opisthosoma usually oval to elongate, cylindrical in M. cyaneotaeniatus , usually with pattern of dark spots dorsally. Male gonopore without epiandrous spigots (examined: M. huanuco , eberhardi, aurantiacus , junin , iguazu : fig. 138, cyaneotaeniatus ). ALS with only one piriform gland spigot each (examined: M. huanuco , eberhardi : figs. 183–184, aurantiacus , junin , iguazu, cyaneotaeniatus ), other spinnerets typical for family.
Sexual dimorphism slight. Females often with more distinct rings on legs, without spines on legs. Epigynum usually with median groove or pocket, sometimes with additional lateral apophyses that are conspicuous in some species.
MONOPHYLY: Most species included share the median groove or pocket on the epigynum (see Specific Relationships for species assigned tentatively).
GENERIC RELATIONSHIPS: The genus is clearly part of the New World clade: male palpal coxa with retrolateral apophysis, epiandrous spigots absent, ALS piriform gland spigots reduced to one, thoracic groove present, exposed tarsal organ, large distance PME-ALE. Otherwise, the phylogenetic relationships are obscure. The cladogram in appendix 2 proposes Coryssocnemis as sister group of Mesabolivar based on the presence of enlarged femora in walking legs. However, this character has considerable homoplasy (see Characters Scored section: char. 23).
SPECIFIC RELATIONSHIPS: The genus can tentatively be divided into four operational species groups; the first three correspond to the core-group mentioned above, the fourth includes species that were assigned to the genus for various reasons, but will probably have to be partly transferred in future more detailed studies. (1) A northern group widely distributed in northern South America, distinguished by the spines on the male metatarsi; including M. eberhardi , huanuco, rubristernus (Caporiacco) , aurantiacus , and cyaneus (Taczanowski) which is possibly a synonym of rubristernus . (2) A closely related group with a similar distribution, without spines on the male metatarsi, with high apophyses on the female epigynum and correspondingly prominent male cheliceral apophyses; including the type species M. pseudoblechroscelis , the closely related M. huambisa , and M. locono , junin, paraensis (Mello-Leitão) , exlineae (Mello-Leitão) . (3) A southern group widely distributed in southern and eastern Brazil and northern Argentina; several species have a distinctively curved procursus, but the group is probably not monophyletic; including M. cyaneomaculatus (Keyserling) , spinulosus, ceruleiventris (Mello-Leitão) , tandilicus (Mello-Leitão) , iguazu, argentinensis (Mello-Leitão) , brasiliensis (Mello-Leitão) , togatus (Keyserling) , guapiara , n. sp., maxacali , n. sp., botocudo, cyaneotaeniatus, azureus (Badcock and Oxon) , and two species whose types I have not been able to examine: M. fluminensis (Mello-Leitão, 1918) , n. comb., and M. nigridentis (Mello-Leitão, 1922) , n. comb. (4) A miscellaneous group, certainly polyphyletic, whose representatives did not fit convincingly into any other genus: M. luteus (Keyserling) , simoni (Moenkhaus) , and possibly also difficilis (Mello-Leitão) , have pockets on the epigynum; M. levii , n. sp., is probably very close to M. luteus ; M. banksi (Moenkhaus) has a curved procursus like that of many representatives of the southern group; M. cambridgei resembles M. huanuco in respect to opisthosoma and epigynum shape, and has spines in bands on the femora like M. spinulosus and M. luteus ; M. xingu , n. sp., is only known from the male and might be an unproblematic species of the southern group.
NATURAL HISTORY: To my knowledge, only M. eberhardi has been studied with some detail (see p. 201).
DISTRIBUTION: From northern South America to northern Argentina. In Colombia the genus is not known from west of the Andes. Together with Metagonia , this is probably the most widely distributed pholcid genus in South America. No representative is known from Central America, the Galápagos Islands, or the Antilles (except Trinidad).
COMPOSITION: The genus as construed here includes 35 nominal species: five species from the northern group (see Specific Relationships above), six species from group 2, 15 species from the southern group, seven species included tentatively, and two species incertae sedis (see below). The collections I have seen contain dozens of further species of this genus from various countries. If the definition is not narrowed, future revisions will probably yield up to hundreds of species. The following species previously included in Blechroscelis are either not congeneric with the type species of Mesabolivar , or are incertae sedis: B. serripes (transferred to Tainonia ), B. modesta (transferred to Waunana ), B. annulipes (transferred to Priscula ). ‘‘ Mesabolivar ’’ globulosus (Nicolet, 1849) and ‘‘ M.’’ aurantius (Mello-Leitão, 1940) are incertae sedis. I have seen what might be the type of B. aurantia from ‘‘Goytacazes,’’ Espírito Santo, in MNRJ (58250); it is a penultimate female. Even though the vial number is correct, it may not be the type, as Mello-Leitão (1940a) describes the epigynum as ‘‘muy alto, bicorne.’’ As for ‘‘ M.’’ globulosus, I do not know if the type material still exists. Mello-Leitão (1941) gave usable illustrations of the epigynum and the male palp, but whether or not his specimens were correctly identified is unknown.
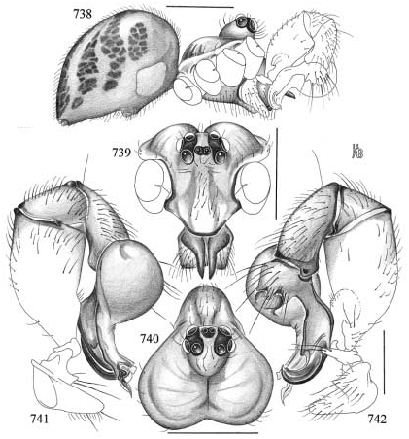
Mesabolivar huambisa , new species Figures 738–747 View Figs View Figs
TYPES: Male holotype, 213 18♀ paratypes from Rio Samiria (4°43'S, 74°18'W), Dept. Loreto, Peru GoogleMaps ; May–June 1990 (T. Erwin ‘‘et al.’’), in MUSM .
ETYMOLOGY: The specific name is a noun in apposition honoring the Huambisa Indians from Peru, whose fierce resistance to subjugation has made them one of the few tribes in South America who still occupy the land that they held in the preconquest era.
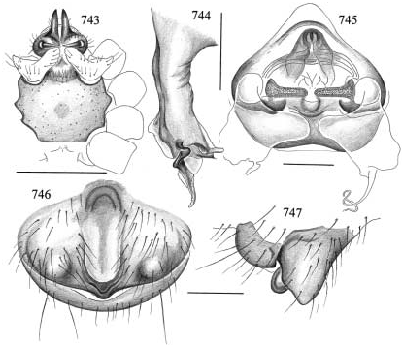
DIAGNOSIS: Closely related to the type species M. pseudoblechroscelis , distinguished by the procursus, which is longer and less strongly bent dorsally in the present species (fig. 742). Distinguished from M. locono by the straight cheliceral apophyses and the shape of the procursus (figs. 738, 742); from M. junin by the shape of the procursus; from M. exlineae and paraensis by the epigynum (shape of lateral apophyses and median pocket, figs. 746–747).
MALE (holotype): Total length 3.0, carapace width 1.3; leg 1: 52.3 (12.7+0.6+12.4 +23.9+2.7), tibia 2: 8.3, tibia 3: 5.7, tibia 4: 8.0; tibia 1 l/d: 93. Habitus as in fig. 738; carapace orange-ochre with light brown spot behind ocular area, with deep thoracic groove; eight eyes on prominently elevated ocular area (figs. 738–739); distance PME- ALE about 65% of PME diameter. Sternum light brown, wide (fig. 743); chelicerae brown proximally, pale ochre-yellow distally, with pair of large straight apophyses, slightly hooked at tips (figs. 738–739). Palp (especially femur) very large in relation to prosoma; coxa with distinct, pointed retrolateral apophysis, trochanter with prominent ventral apophysis (fig. 742), femur proximally with retrolateral apophysis (fig. 742); procursus curved distally (fig. 742); bulb as in figs. 741, 744, with slightly spiraling apophysis. Legs ochre to light brown, femora and tibiae with light tips; legs without spines, without curved and vertical hairs; retrolateral trichobothrium of tibia 1 at 3%; tarsus 1 with over 30 pseudosegments. Opisthosoma pale greenish, with darker greenish spots dorsally.
VARIATION: Tibia 1 in 6 male paratypes: 10.7–13.6 (x¯ = 11.8). In the male from Napo, Alinahui, Ecuador, the tip of the procursus is slightly different ; the male and the female from this locality are therefore assigned tentatively.
FEMALE (paratypes): Tibia 1 (N = 5) 7.9– 8.8 (x¯ = 8.3). In general very similar to male, but with distinct dark rings on femora (subdistally) and tibiae (proximally and subdistally), subdistal rings followed by light tips. Epigynum dark brown, with median pocket and pair of lateral apophyses (figs. 746–747); internal genitalia with distinctively formed pore plates, and large membranous structures of unknown function reaching into lateral apophyses (fig. 745).
DISTRIBUTION: Known from northern Peru and Ecuador.
MATERIAL EXAMINED: PERU: Loreto: Rio Samiria : types above ; same locality, June 16, 1990 (T. Erwin ‘‘et al.’’), 33 in MUSM ; Cocha Shinguito (5°08'S, 74°45'W), June 18, 1990 (T. Erwin ‘‘et al.’’), 13 in MUSM. EC- UADOR: Napo: 20 km E Puerto Napo, Alinahui (1°00'S, 77°25'W), 450 m elev., Jan. 1994 (V. D. & B. Roth), 13 assigned tentatively, in CAS GoogleMaps ; same data, in building, 1♀ assigned tentatively, in CAS GoogleMaps ; Reserva Faunistica Cuyabeno, Laguna Grande (0°00'S, 76°10'W), from Macrolobium trees in lake, June 28, 1988 (W. Maddison), 13 2♀ (2 vials) in MCZ GoogleMaps ; Pompeya, Napo River , May 1965 (L. Peña), 13 1 juvenile in MCZ .
Mesabolivar locono , new species Figures 748–755 View Figs
TYPE: Male holotype from Lawa River , Benzdorp, Marowijne Dist., Surinam ; ‘‘forest night sweep,’’ Nov. 6, 1963 (B. Malkin), in AMNH .
NOTE: It is possible that this is a junior synonym of M. paraensis (of which only the female is known; see below).
ETYMOLOGY: The species name is a noun in apposition, honoring the Locono people of eastern Venezuela, Guyana, Surinam, and French Guiana.
DIAGNOSIS: Distinguished from M. huambisa by the curved cheliceral apophyses (fig. 750); from M. junin and M. pseudoblechroscelis also by the shape of the procursus (figs. 753–755).
MALE (holotype): Carapace width 1.4, carapace length 1.1 (opisthosoma missing); leg 1: 34.0 (8.3+0.3+8.4+15.1+1.9), tibiae 2 and 3 missing, tibia 4: 5.7; tibia 1 l/d: 79. Colors poorly preserved (entire animal light brown); carapace with deep thoracic groove; eight eyes on prominent elevation (fig. 748); distance PME-ALE about 65% of PME diameter. Chelicerae with pair of large, strongly curved apophyses (fig. 750). Palp (especially femur) very large in relation to pro- soma; coxa with distinct apophysis, femur proximally with retrolateral apophysis and small ventral hump slightly more distally (fig. 753); procursus rather simple, strongly curved (figs. 754–755); bulb relatively simple (globular part shrunken in type) (fig. 752). Legs apparently without spines, without curved and vertical hairs (many hairs missing). Opisthosoma missing.
VARIATION: The male from Guyana (see below) differs minimally with respect to the tip of the procursus and is slightly larger (tibia 4: 6.5), but otherwise indistinguishable.
FEMALE: Unknown (see Note above).
DISTRIBUTION: Known from Surinam and Guyana.
MATERIAL EXAMINED: SURINAM: Marowijne: Benzdorp: type above. GUYANA: Mazaruni-Potaro: Kartabo Point, Dec. 22– 24, 1983 (W. Steiner, J. Byrd, J. Hill, F. Holtzclaw), 13 in USNM.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |