
Melitaea mimetica Higgins, 1940
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.830.1865 |
|
publication LSID |
lsid:zoobank.org:pub:C1F47AD9-ECF6-4F0F-9928-55A45332FF4B |
|
DOI |
https://doi.org/10.5281/zenodo.6855844 |
|
persistent identifier |
https://treatment.plazi.org/id/EB42F330-FFDD-B32D-9149-FE8AFD5CF99E |
|
treatment provided by |
Felipe |
|
scientific name |
Melitaea mimetica Higgins, 1940 |
| status |
|
Melitaea mimetica Higgins, 1940 View in CoL
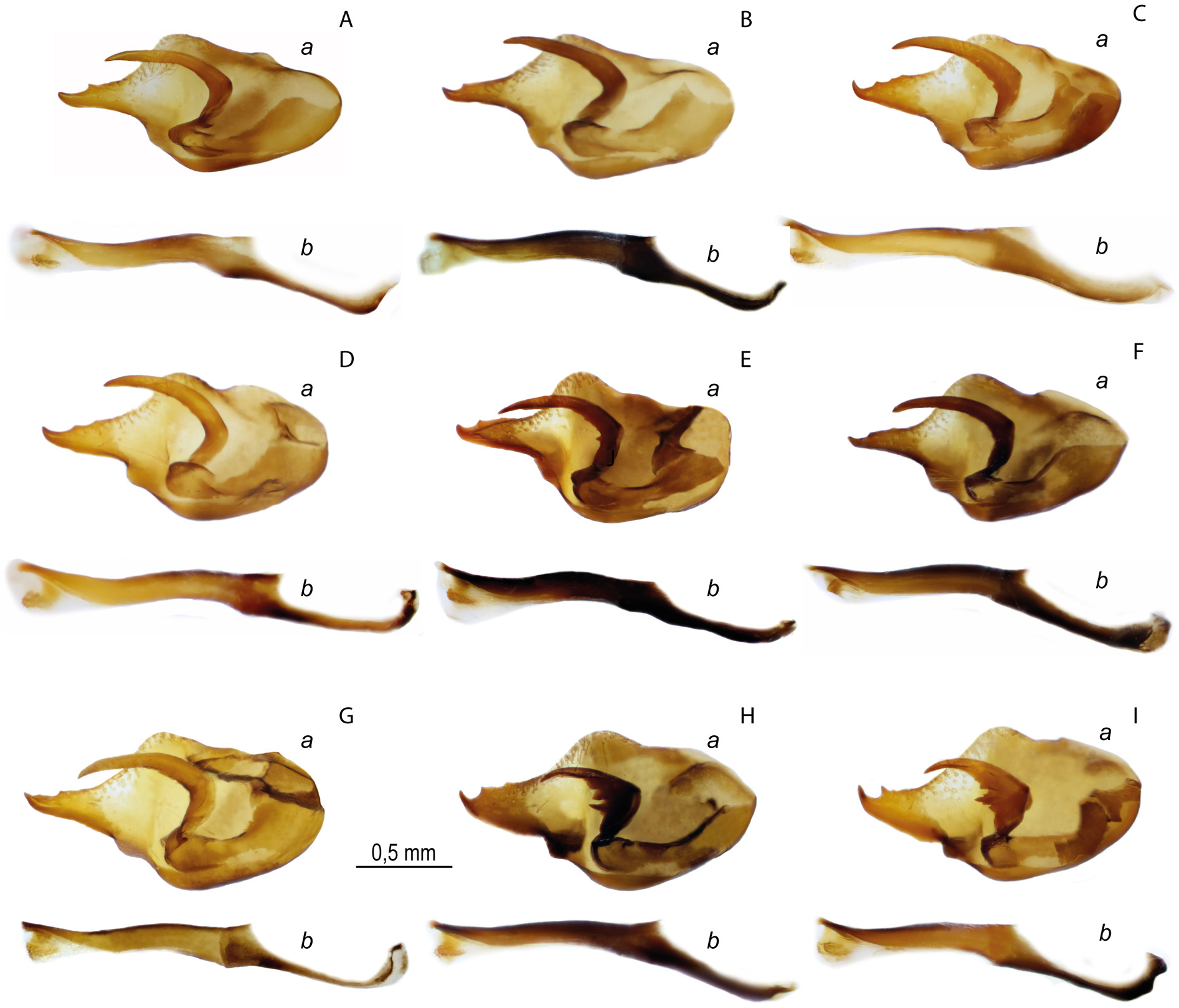
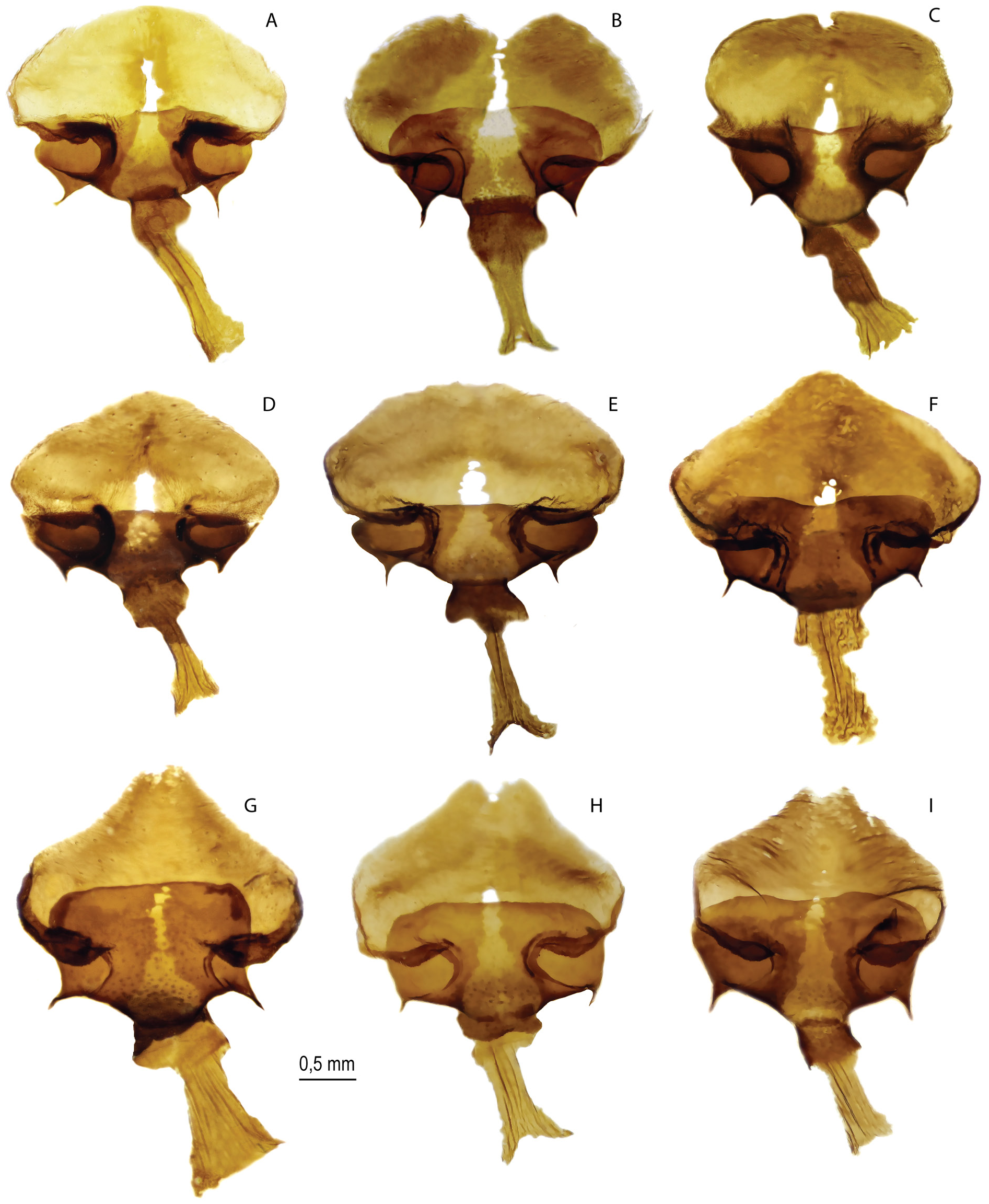
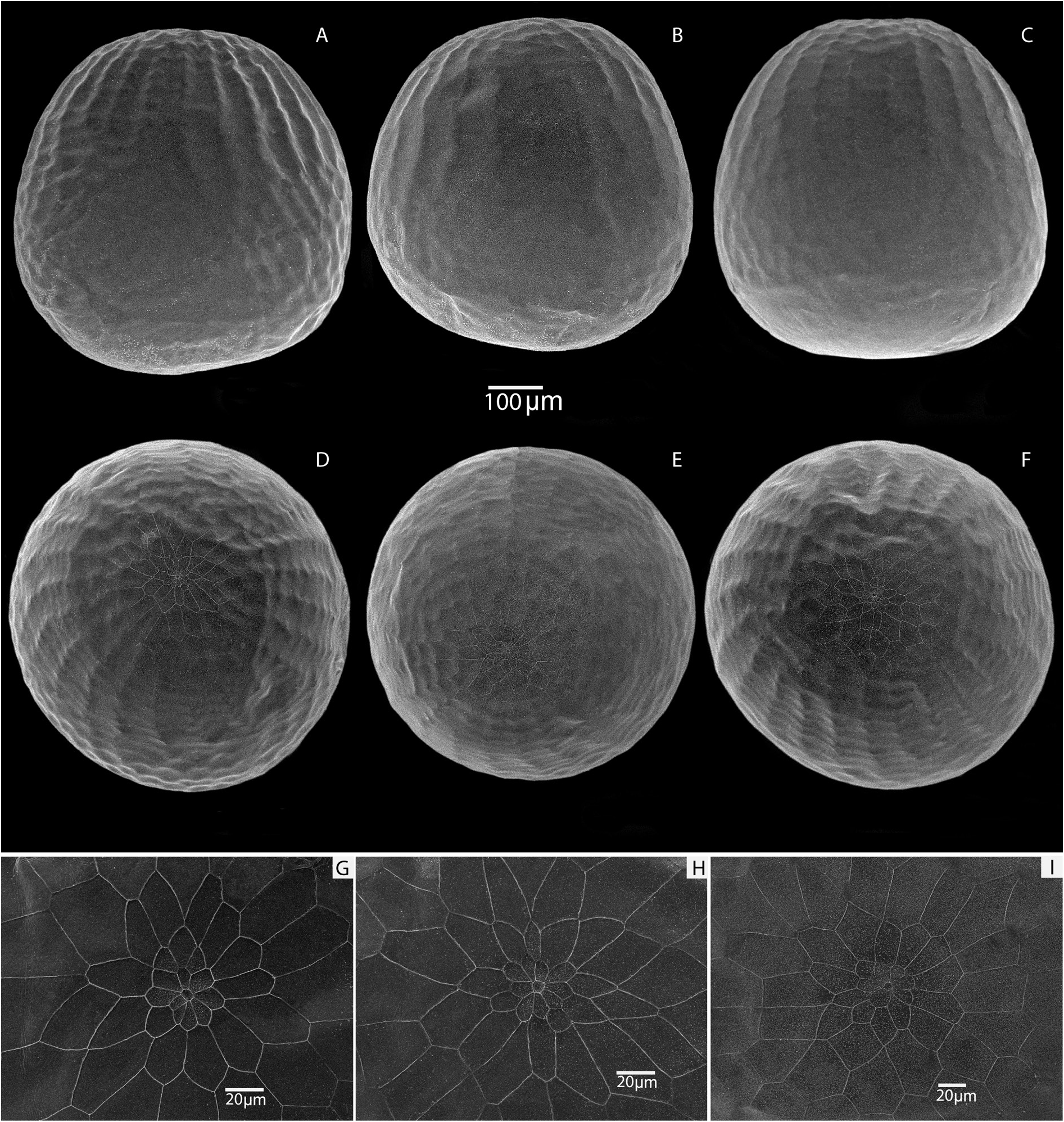
Figs 1C–D View Fig , 3A – G View Fig , 4F – I View Fig , 6–8 View Fig View Fig View Fig , 16D – H View Fig , 18F – I View Fig , 29C, E View Fig , 30 View Fig ; Table 1 View Table 1
Melitaea lukto [sic] mimetica Higgins, 1940: 52 .
Type locality: “ Balochistan, Khojak” [ Pakistan, Balochistan, Khojak Pass].
“ Melitaea deleréi View in CoL nov. spec. ” Heydemann, 1954: 415, pl. 35 fig. 9 (holotype ♀), pl. 35 fig. 8 (allotype ♂), fig. 10 [genitalia].
Type locality: “ Kabul Umgebung” [ Afghanistan, Kabul vicinity].
Remarks
The taxon delerei was described from two specimens: 1 ♀ (holotype) and 1 ♂ (allotype). The butterflies were collected in the vicinity of Kabul on 23 May 1951 and 28 Apr. 1953, respectively. The text contains photos of the holotype ♀ and the allotype ♂ ( Heydemann 1954). The author of the original description does not mention the existence or number of other type specimens. Van Oorschot & Coutsis (2014) provide other data on the type material of delerei , and these are quoted here without changes: “Type material: ♂ holotype, 4 ♂♂, 2 ♀♀ paratypes (coll. Görgner, including ♂ with yellow label stating “Kabul, ♂, 23.5.1951, leg Delere, Heydemann, Kiel”, ♂ with orange label stating “ ♂, Mel. dodgsoni delerei, Hdm. ”, ♂ with red label stating “male, Holotypus Mel. delerei Hdm. 11.54.)”. The type material is stored in the Ernst Görgner collection (Cosswig Anhalt, Germany). We do not know the reason for such a serious discrepancy in the data on the type material of the taxon delerei .
Type material
The taxon mimetica is described based on the holotype, allotype, and 10 paratypes from Balochistan ( Pakistan) from the following localities: Khojak, Ziarat Rd., Zaghun, Gawar, Sheik Wazil, Urak and Quetta ( Higgins 1940). The number and the gender of specimens for each collection point are not specified. According to Smith (1988) the type material stored in the BMNH consists of 3 ♂♂, 1 ♀ with the labels “ Pakistan, Zaghun, 6000 ft. ” and 1 ♀ with the label “Gawar”. The storage location of the type specimens from Sheik Wasil, Urak and Quetta is unknown.
The type locality of the holotype is mentioned by Higgins as Khojak Pass , and the place of capture of the allotype is mentioned as “ Ziarat Rd. ” ( Higgins 1940). However, in the work devoted to the bibliography and catalog of type specimens of taxa described by Higgins ( Smith 1988), it is indicated that the specimen marked by Higgins as an allotype has the label “Khojak”, and the holotype, on the contrary, has the label “Ziarat Rd.” In accordance with recommendation 76A.1.3. (ICZN 2004), the holotype should be considered a specimen caught on the Khojak Pass.
Paratype ( Fig. 1C–D View Fig ) We have photos of the paratype (allotype), ♀, with the following labels: “Holotype [!]” (printed label); “ Ziarat Rd. / Balochistan, 6000 w. / 20.04.31” (handwritten label); “Roth. B.-1” (printed label); “ Melitaea lutko mimetica / type ♂, L. G. Higgins ” (handwritten label) ; “ Melitaea lutko / mimetica Higgins / C. R. Smith det. 1986, Holotype (!)” (handwritten + printed label); BMNH .
Notes on diagnosis
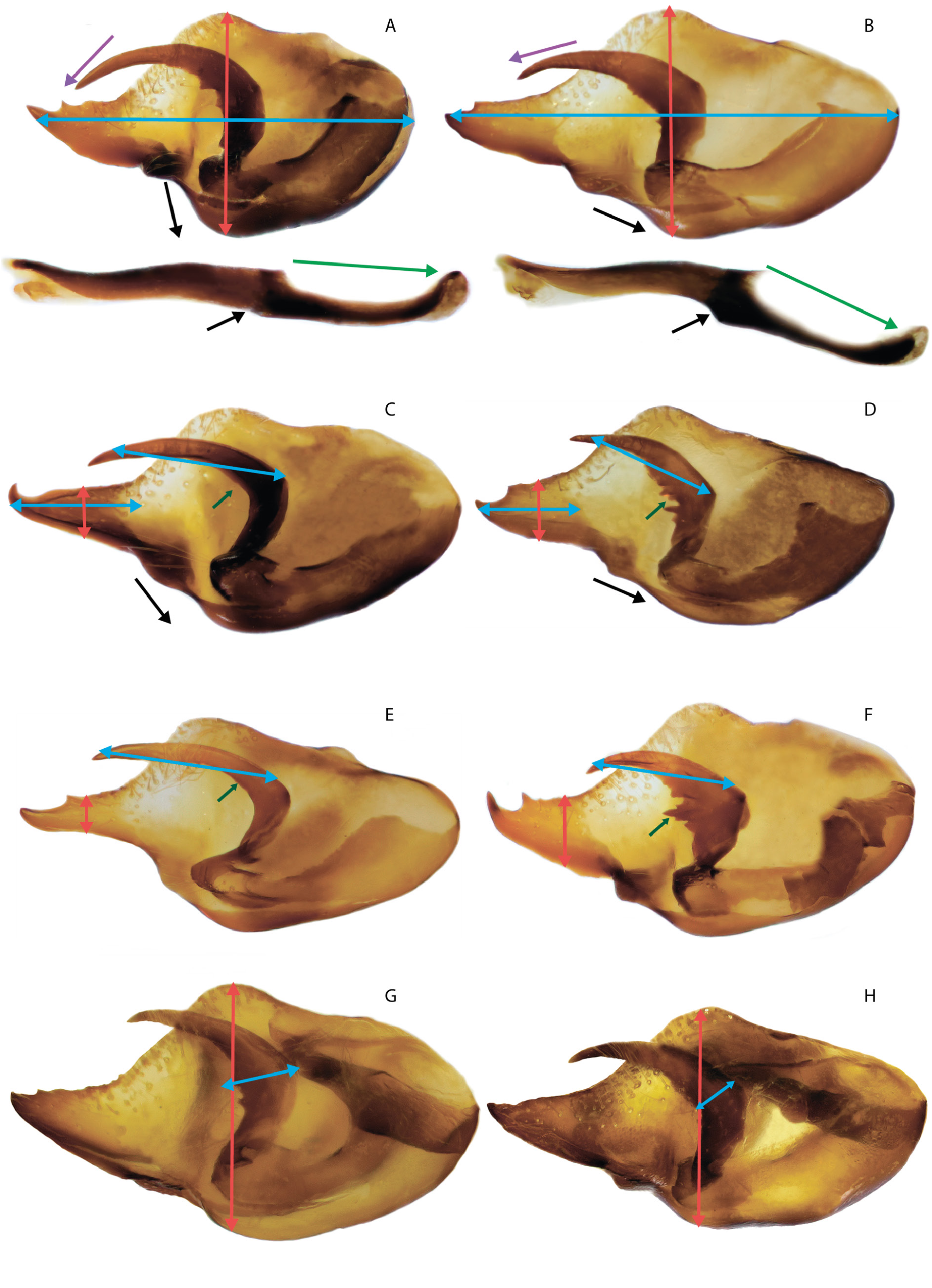
The taxon delerei was considered as a synonym of the taxon mimetica based on external features and the structure of the genitalia ( Higgins & Wiltshire 1956), or together with the taxon mimetica as a synonym of M. lutko ( van Oorschot & Coutsis 2014) . In the latter case, they were able to study the genital apparatus of the paratype of the taxon delerei , whose structure in their opinion is transitional between M. lutko and M. timandra . However, the description of M. delerei clearly states that one of the features of the male is a very long and thin harpe without teeth on the inner side, which is a characteristic feature of M. mimetica . In general, the genital apparatus of the male delerei , an image of which is given by Heydemann (1954), is characterized by a thin caudal process of the valva with 2–3 teeth located distally on the dorsal side and a long braid-shaped harpe. Judging from the image, the male and female have a well-expressed postdiscal pale area on the UPF. In this case, we agree with Higgins’ opinion and believe that the taxon delerei has all the distinctive features of M. mimetica and should be treated with the latter as a synonym. We do not exclude the subspecies status of the taxon delerei , but a final decision can be made only after the identification and study of the holotype and additional material from Afghanistan.
Material examined
PAKISTAN – Balochistan • 2 ♂♂ (all dissected), 2 ♀♀ (all dissected); Quetta, Urak ; alt. 2400–2700 m; 10–14 May 1983; Eckweiler leg.; EDMSU • 1 ♂; Ziarat ; alt. 2400–2700 m; 17–23 May 1983; Eckweiler leg.; EDMSU .
AFGHANISTAN – Bamian Prov. • 6 ♂♂ (5 dissected); 10 km S of Bamian t., Hushkak v. vicinity ; alt. 2700–2800 m; 2 Jun. 2012; O. Pak leg.; EDMSU • 7 ♂♂ (5 dissected), 3 ♀♀ (all dissected); Punjub Distr. , 10 km NE of Varas v.; alt. 2400 m; 20 may 2012; I. Pljushtch leg.; EDMSU • 1 ♀ (dissected); 8 km S of Bamian, Koh-e-Baba Mts, Dara-e-Khushkak ; alt. 2930 m; 12 Jun. 2016; I. Pljustsh leg.; EDMSU • 7 ♂♂; Panjub Distr. ; 10 km NE of Varas vil.; alt. 2400 m; 21 May 2012; I. Pljushtch leg.; coll. I. Pljushtch • 13 ♂♂, 5 ♀♀; Panjub Distr., Varas vil.; alt. 2400 m; 21 May 2012; I. Pljushtch leg.; coll. I. Pljushtch. – Ghor Prov. • 7 ♂♂ (5 dissected), 2 ♀♀ (1 dissected); 17 km E of Changcharan, 15 km S of Bandi-Ali, Gazak Mts ; alt. 2400 m; 26 May 2012; O. Pak leg; EDMSU • 3 ♂♂ (all dissected), 1 ♀ (dissected); 16 km E of Changcharan, Bandi-Ali v. vicinity ; alt. 2400 m; 26 May 2012; I. Pljushtch leg.; EDMSU • 1 ♂ (dissected), 1 ♀ (dissected); Bayan Ridge, 15 km S of Changcharan, Kindival valley ; alt. 2800 m, 27 May 2012; I. Pljushtch leg.; EDMSU • 1 ♂; Bayan Ridge, Changcharan circ., Kindaval valley ; alt. 2800 m; 27 May 2012; I. Pljushtch leg.; coll. I. Pljushtch • 4 ♂♂; 16 km E of Changcharan, Bandi-Ali vil.; alt. 2400 m; 26 May 2012; I. Pljushtch leg.; coll. I. Pljushtch. – Kabul Prov. • 1 ♀; Gargha ; alt. 2000 m; 1 Jun. 2010; I. Pljushtch leg.; coll. I. Pljushtch.
Redescription
Male ( Fig. 6A–D, I–L View Fig )
WINGS. FW length is 17.5–19 mm, the paratype is 18 mm. UPS ground color is yellow-orange or pale orange-red; UPS black marginal border is thin with well-defined marginal spots along the outer edge of the wings; UPS submarginal row represented by small pointed black lunules; UPF discal row is formed by expanded black spots fused with each other near the costa; UPF postdiscal pale-yellowish area, located behind the black discal spots, is well expressed along the entire length and contrasts with the general background. There is a pale-yellowish spot in the distal part of the discoidal cell; UPH discal row is usually reduced; UPH basal suffusion covers no more than ⅓ of the surface of the wing. UNF ground color is pale orange-red, with a well-defined pale area of the outer edge of the wing between the veins Sc and Cu1 and in postdiscal area between veins Sc and M1. UNH ground color is white without the admixture of dark scales. UNH lunules forming the proximal edge of submarginal orange fascia outwardly concave and sharply pointed between veins M3 and Cu2.
MALE GENITALIA ( FigS 3A–G View Fig , 16D–H View Fig , 18F–I View Fig , 29C, E View Fig ). The valva is oval with a slender caudal process, usually with one small spine (or without it) on the dorsal surface in the distal part. The long harpe is narrow, without spines (or in rare cases with one or two weakly developed spines) on the inner side. The aedeagus is curved in the central part, distally with a rounded convex dorsal edge. The posterior part of the aedeagus is mostly located at an angle to the anterior part and is directed downward. There is often a well-marked protrusion on the ventral side at the junction of the two parts of the aedeagus. The wide saccus is rounded distally, its length being 1.5 times greater than the width.
Female ( Fig. 6E–H, M–P View Fig )
WINGS. FW length is 20–21 mm. UPS ground color is pale orange-red. UPF postdiscal pale area is well expressed and represented by whitish-yellow spots fused to the costal edge and contrasting well with the wing ground color. UPF well-marked pale macule is present in the discoidal cell. UPF submarginal row on the is represented by dark pointed spots, and on the UPH by thin black lunules. UPH black distal row is absent or weakly expressed. UNH pattern is similar to that of males.
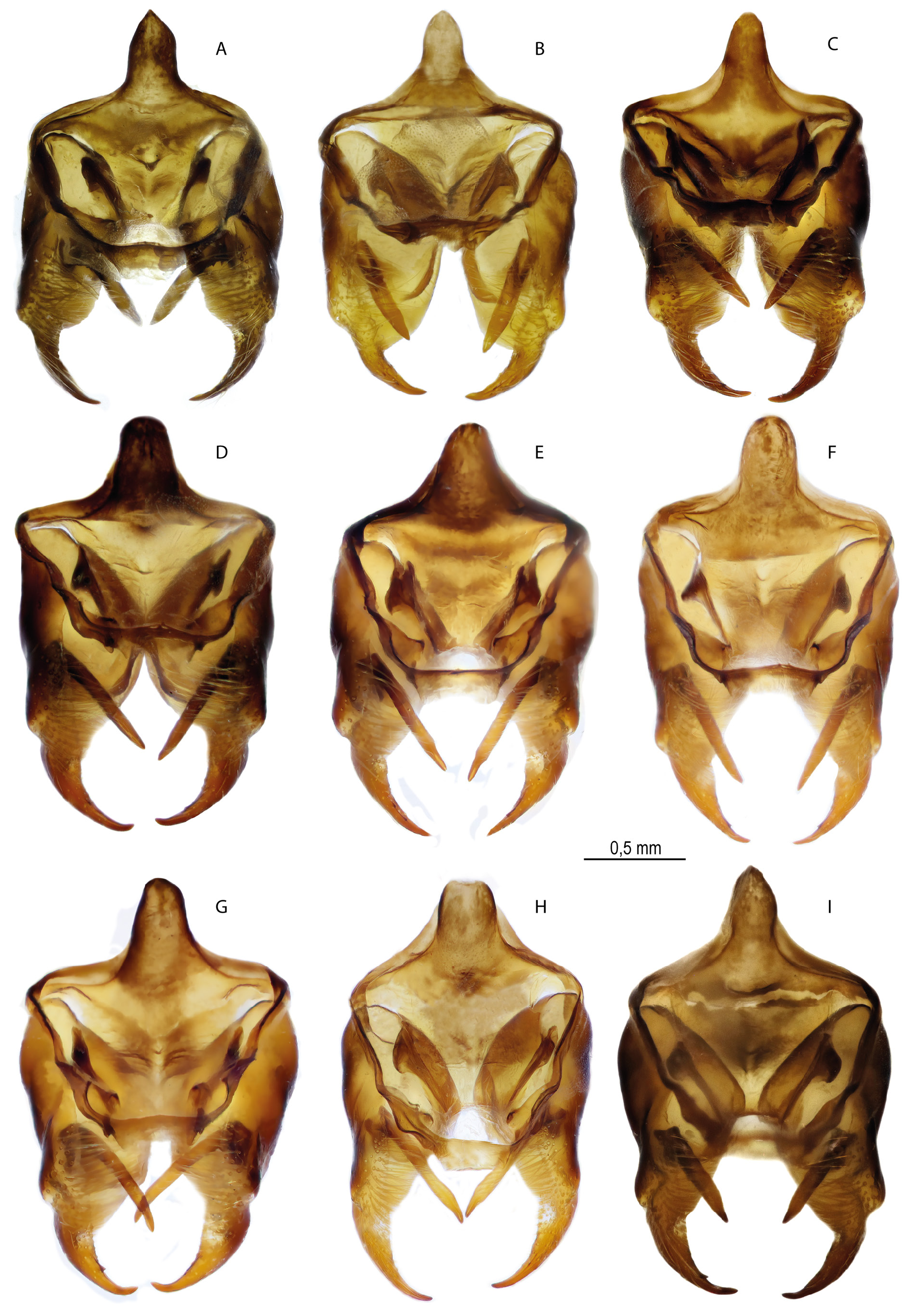
FEMALE GENITALIA ( Fig. 4F–I View Fig ). The postvaginal plate is rounded-trapezoidal in shape. The antevaginal plate is expanded in the dorsoventral direction, its outer edge noticeably extends beyond the boundaries of the bend of the postvaginal plate (auricules).
Preimaginal stages: eggs ( Fig. 7 View Fig , Table 1 View Table 1 )
Material examined: 10 eggs from 1 ♀; Afghanistan, Ghor Prov., 16 km E of Changcharan, Bandi-Ali v. vicinity , alt. 2400 m.
The egg is barrel-shaped. The height of the eggs is from 624.0 µm to 626.6 µm, the width is from 598.0 µm to 603.5 µm. The sculpture of the micropile region is formed from four to five rows of pentahexagonal cells of various lengths and widths. The diameter of the micropile rosette in the widest part varies from 37 µm to 41 µm. The micropile rosette is formed by 8–9 primary quatro-pentahedral cells 5.0 µm to 16.0 µm wide and 9.0 µm to 23.0 µm long. The micropile is rounded in shape with an average diameter of 6.6 µm. There are 23–24 lateral longitudinal ribs that limit the micropile area and decrease to ⅓ of the egg surface. The transverse ribs are weakly expressed. Below the lateral ribs the chorion is relatively smooth.
A distinctive feature of the morphology of eggs of M. mimetica is their well-defined barrel shape. In addition, M. mimetica has the smallest eggs among the representatives of the lutko group. Their maximum height is 626.6 µm. However, the egg of M. timandra , which is oval with a slight narrowing in the apical region ( Kolesnichenko & Kotlobay 2020), has a height of about 850 µm, whereas that of M. shahvarica sp. nov., whose eggs are pear-shaped, has an average height of about 700 µm. In addition, the M. mimetica micropile rosette is formed by 8–9 primary cells, while in the M. timandra egg the micropile rosette is formed by 9–10 cells, and in the M. shahvarica egg the micropile is surrounded by 7–8 primary cells.
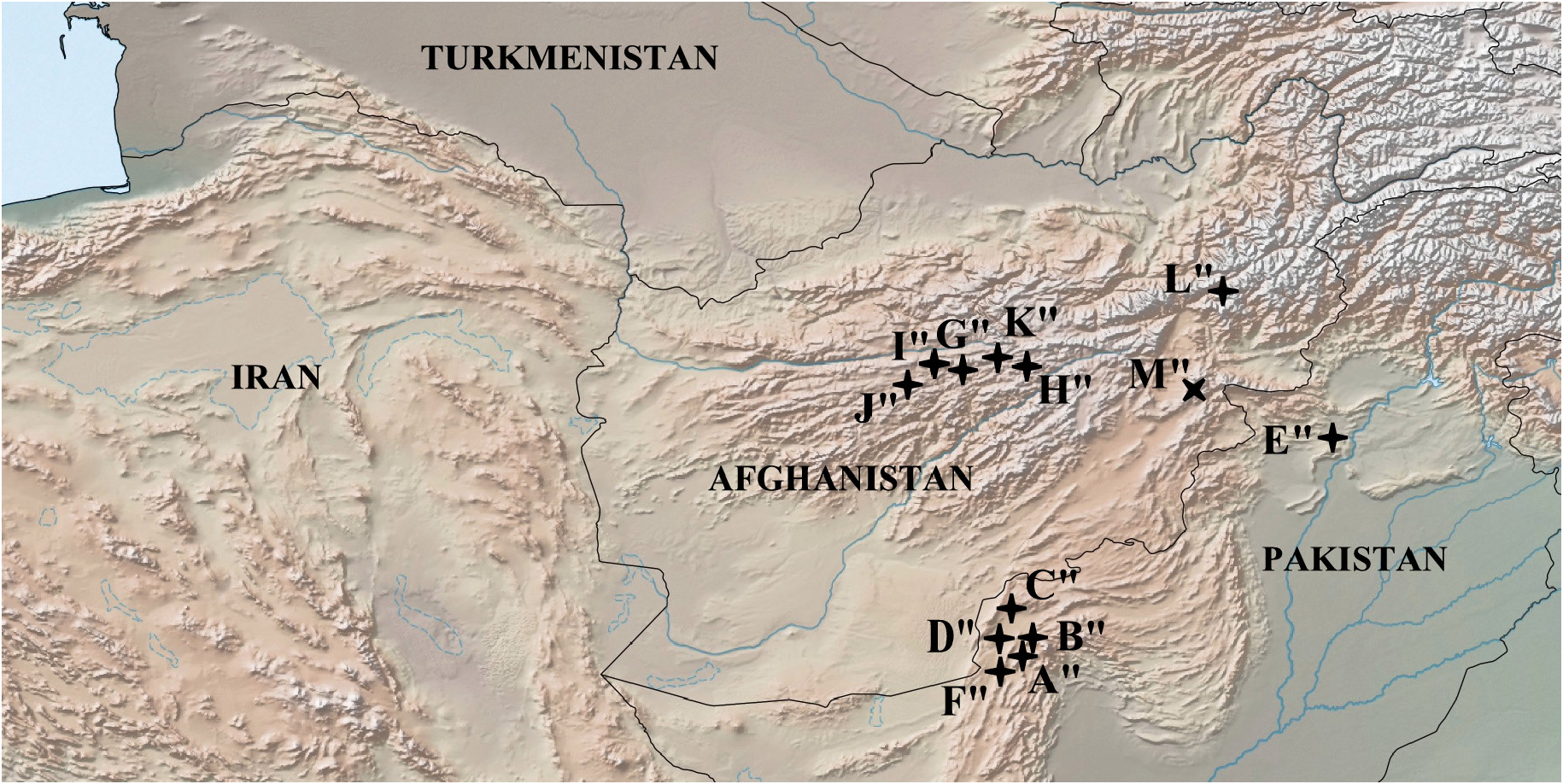
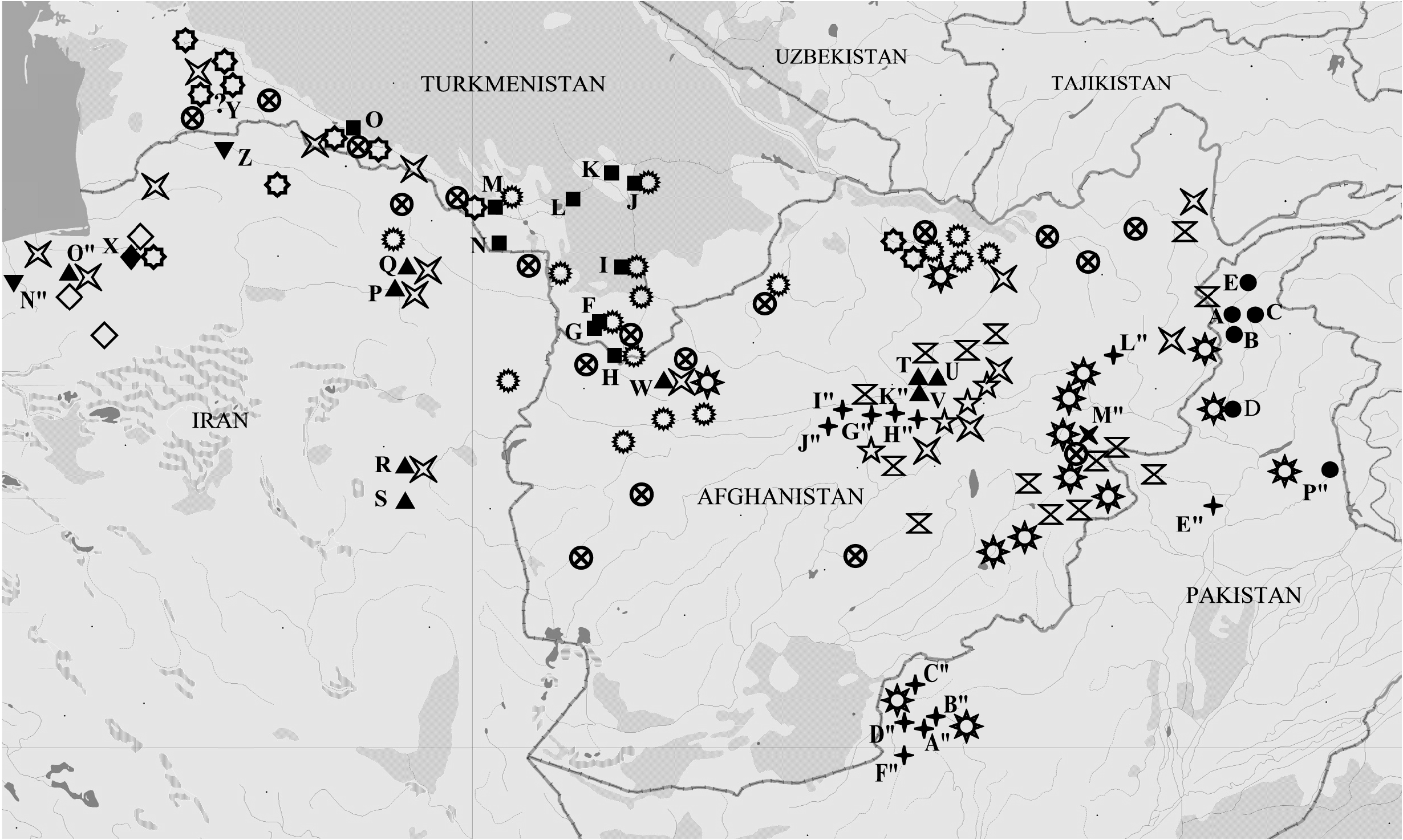
Distribution ( Fig. 8 View Fig )
Pakistan: Balochistan, Punjab; Afghanistan: Panjshir Gorge, Koh-i-Baba Ridge, mountains in the vicinity of Punjab, Bandi-Bayan Ridge.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Nymphalinae |
|
Genus |
Melitaea mimetica Higgins, 1940
| Kolesnichenko, Kirill A. & Kotlobay, Anatoly A. 2022 |
Melitaea lukto [sic] mimetica
| Higgins L. G. 1940: 52 |