
Medusapyga alsea LaBonte, 2023
|
publication ID |
https://doi.org/ 10.5281/zenodo.11067222 |
|
DOI |
https://doi.org/10.5281/zenodo.12613601 |
|
persistent identifier |
https://treatment.plazi.org/id/51533818-FFE3-4765-FF85-BF1FFBCEF9D0 |
|
treatment provided by |
Felipe |
|
scientific name |
Medusapyga alsea LaBonte |
| status |
sp. nov. |
Medusapyga alsea LaBonte , new species
Holotype. Male, deposited in the California Academy of Sciences, labeled: “ OREGON, Benton Co., Prairie Peak, 11 km S Alsea, J.R. LaBonte, 9 V 1993, elev. 810 m, 44.2844°N 123.5858°W, under stones”, “HOLOTYPE, Medusapyga alsea LaBonte sp. nov., designated 2022 [red paper]”. GoogleMaps
Paratypes. 329 (258 females, 71 males), residing in the following collections and institutions: CAS (32), CMNH (2) , CNC (2) , EMEC (1) , GLPC (84) , JRLC (155) , MCZ (2) , MNHN (2) , NHMUK (2) , NMNH (2) , ODAC (17) , OSAC (22) , SDSU (4) , WSU (2) . USA, Oregon, Benton Co., Prairie Peak, 11 km south of Alsea, 44.2844°N 123.5858°W, 810 m elevation [Type locality] (296). 9 V 1993, G.L. Peters (63), J. R. LaBonte (57) GoogleMaps . 14 V 1993, J. R. LaBonte (10) . 19 VI 1993, G.L. Peters (18), J. R. LaBonte (19) . 26 VI 1993, G.L. Peters (1) . 31 VII 1993 , J. R. LaBonte, (10) . 7 V 1994, G.L. Peters (2) , J. R. LaBonte (7) . 15 V 1995, J. R. LaBonte (6) . 6 V 1998, J. R. LaBonte (6), R.L. Westcott (6). 29 IV 2002, D.H. Kavanaugh (21), J. R. LaBonte (13) . 1 V 2003, J. R. LaBonte and B. Smith (12), R.L. Westcott (9) . 20 III 2010, D. R. Maddison & J. R. LaBonte (1) . 29 IV 2014, D. H. Kavanaugh and J. R. LaBonte (1), J. R. LaBonte (1). 29 IV – 9 XI 2014, soil trap, J. R. LaBonte (27) . 2 IV 2022, D. R. Maddison & P. R. Triplett (1) . 15 V 2022, D. R. Maddison (5). GoogleMaps USA, Oregon, Benton Co., Prairie Peak, 11 km south of Alsea , 0.6 km west of the type locality, 44.2848°N 123.5933°W, 750 m elevation (32). 6 V 1998, R.L. Westcott (2) GoogleMaps . 29 IV 2002, D.H. Kavanaugh (7), J. R. LaBonte (4) . 1 VI 2004, J. R. LaBonte (9). 20 III 2010, D. R. Maddison & J. R. LaBonte (1). 30 V 2010, D. R. Maddison (4). 29 IV – 9 XI 2014, soil trap, J. R. LaBonte (2). 2 IV 2022, D. R. Maddison & P. R. Triplett (1). GoogleMaps 15 V 2022, D. R. Maddison (1). GoogleMaps 19 VI 2022, K.W. Will (1). GoogleMaps USA, Oregon, Benton Co., Prairie Peak, 10 km south of Alsea , 1.1 km WNW of the type locality, 44.2881°N 123.5982°W, 715 m elevation (1). 15 V 2022, L.A. Martin (1). GoogleMaps
Depending upon the GPS system used, and the precise spot the measurements were taken within each locality, the exact geographic coordinates and elevations may slightly differ among the labels of the different collectors and institutions.
Type Locality. USA, Oregon, Benton Co., Prairie Peak, 11 km south of Alsea , 44.2844°N 123.5858°W, 810 m elevation GoogleMaps .
Etymology. The specific epithet refers to the Oregon Coast Range river drainage of this name and the community nearest the type locality. The name is to be treated as a noun in apposition.
Description. Habitus ( Fig. 2A View FIGURE ). SBL 2.20-2.65 mm. Color translucent dark testaceous to reddish brown to dark brown. Convex in lateral view.
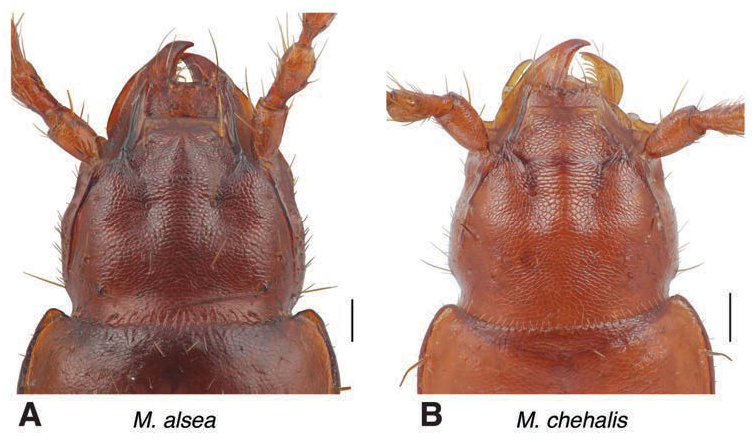
Head ( Figs 1A View FIGURE , 7A View FIGURE ). The medial convexity formed by the frontolateral furrows ends in an abrupt declivous prominence near the anterior margin of the clypeus. In dorsal view, it is roughly triangular, pointed at the anterior extreme and broadening posteriorly. In lateral view, it ends at the anterior in a steep precipice. Tempora shallowly convex, rounded.
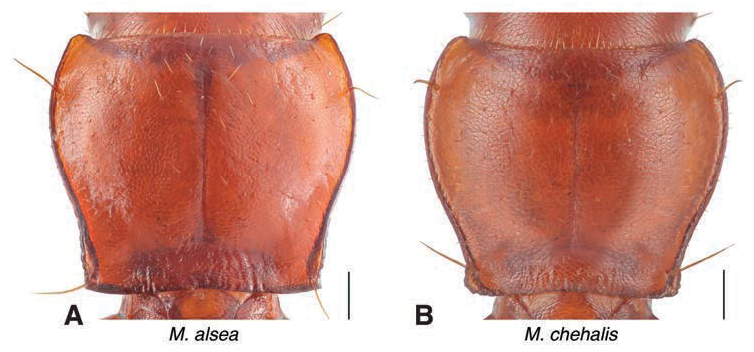
Pronotum ( Fig. 8A View FIGURE ). PWm/ PL ~1.15, PWa and PWp approximately equal. Widest at about anterior third. Anterior angles strongly protuberant. Anterior margin truncate. Lateral margins distinctly sinuate, approximately parallel from posterior angles to well anterior of posteromedian impression, obliquely arcuate to PWm, then convergently arcuate to anterior angles. Lateral explanations distinct and reflexed, widest at anterior and posterior angles, narrowing from medial posterior impression anteriorly to just posterior of anterior angles, the point at which the explanations are narrowest. Lateral margins with fine, distinct denticles where margins are parallel anterior of hind angles but not extending to posterior margins of hind angles. Posterior angles right to slightly acute, sharp. Disc strongly convex. One long fixed stout seta just anterior of each posterior angle and a longer fixed seta located laterally at approximately the anterior quarter of each side.
Elytra ( Figs 2A View FIGURE , 9 View FIGURE ). Greatest width just anterior of middle. Convex in lateral view, rather steeply ascending from anterior to just before middle, more gradually descending to apex, more or less evenly convex in cross section. Each elytron with lateral margin coarsely denticulate from humerus to about anterior one-fourth, thereafter finely denticulate to plical crossing. Humeri truncate anteriorly, distinctly and obtusely angulate. Lateral margins slightly obliquely divergent in anterior third, thereafter evenly convergently arcuate to the plical crossing, shallowly indented there, and then obliquely or obliquely-arcuate to the separately rounded apices. Intervals slightly convex.
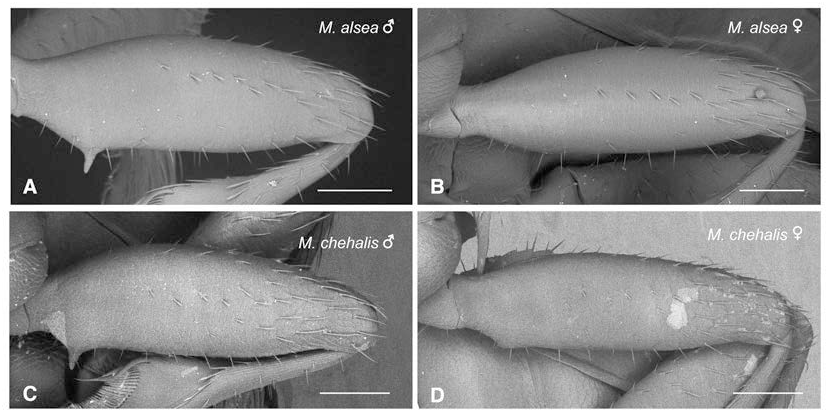
Legs. Profemoral spine of males near basal one-quarter of ventral face, with its length about three times its greatest diameter ( Fig. 4A View FIGURE ). Females lack any evidence of the profemoral spine found in males or the carina that in males extends from the profemoral base to the male profemoral spine; the base of the ventral face of the female profemur is thus evenly rounded ( Fig. 4B View FIGURE ). Mesotrochanter with large, blunt, triangular tooth projecting from posteromedial margin ( Fig. 13A View FIGURE ).
Abdominal ventrites. Apex of intercoxal process of second visible abdomnal ventrite acute. A small fovea is present on the second visible abdominal ventrite just posterior of the intercoxal process between the metacoxae. It is the shape of an elongate oval, narrowest anteriorly and broadest near the posterior margin; it is thus tear-drop shaped. The surface of the fovea is smooth except for microsculpture.
Male genitalia ( Fig. 11A,B View FIGURE ). Aedeagus arcuate, apex angled downward; internal sac when viewed from left side with Cshaped structure.
Sexual dimorphism. In addition to the differences in protarsomeres and profemora, females are larger than males. Female SBL ranged from 2.40– 2.65 mm and averaged 2.50 mm. Male SBL ranged from 2.20– 2.48 mm and averaged 2.38 mm. A two-tailed Student’s T-test indicated these differences in average length were highly significant (p = 0.000001). The medial anterior fovea posterior to the intercoxal process of visible ventrite 2 appears slightly larger and deeper in females.
Variation. The posterolateral pronotal margins of some specimens were slightly obliquely divergent, rather than parallel. Some individuals were distinctly paler than others, with the most extreme examples a very pale yellowish. The latter may have been teneral individuals.
Identification. Characters of M. alsea distinguishing it from M. chehalis include greater average and overall length, more robust and convex habitus, a frontoclypeal prominence, pronotum with protruding anterior angles and sharply rectangular or acute posterior angles, mesotrochanter with large triangular tooth at middle of the posterior margin, the apunctate surface of the abdominal fovea, the absence of a keel on visible abdominal ventrite 2 of males, and the more arcuate aedeagus with a C-shaped structure in the internal sac.
Geographic distribution ( Fig. 15 View FIGURE ). Medusapyga alsea is known only from three closely approximate sites in central western Oregon: the type locality, 44.28435°N 123.58578 ° W, at 810 m elevation; a site 0.6 km W of the type locality, 44.28481°N 123.59331 ° W, at 750 m elevation; a site 1.1 km WNW of the type locality, 44.2881°N 123.5982°W, at 715 m elevation.
Note. This species was referred to as “ Anillina ‘USA: Oregon’” in Maddison et al. (2019).






| CAS |
USA, California, San Francisco, California Academy of Sciences |
| CMNH |
USA, Pennsylvania, Pittsburgh, Carnegie Museum of Natural History |
| OSAC |
USA, Oregon, Corvallis, Oregon State University |
| CAS |
California Academy of Sciences |
| CMNH |
The Cleveland Museum of Natural History |
| CNC |
Canadian National Collection of Insects, Arachnids, and Nematodes |
| EMEC |
Essig Museum of Entomology |
| MCZ |
Museum of Comparative Zoology |
| MNHN |
Museum National d'Histoire Naturelle |
| NHMUK |
Natural History Museum, London |
| ODAC |
Oregon Department of Agriculture |
| OSAC |
Oregon State Arthropod Collection |
| SDSU |
Severin-McDaniel Insect Collection |
| WSU |
Weber State University, Bird and Mammal Collection |
| R |
Departamento de Geologia, Universidad de Chile |
| V |
Royal British Columbia Museum - Herbarium |
| VI |
Mykotektet, National Veterinary Institute |
| PL |
Západoceské muzeum v Plzni |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Trechinae |
|
Genus |