
Martinidrilus carolinensis, Fend, Steven V. & Lenat, David R., 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.180092 |
|
DOI |
https://doi.org/10.5281/zenodo.6252652 |
|
persistent identifier |
https://treatment.plazi.org/id/03953B40-FFBC-FFFA-FF35-E35FFB40CD48 |
|
treatment provided by |
Plazi |
|
scientific name |
Martinidrilus carolinensis |
| status |
sp. nov. |
Martinidrilus carolinensis View in CoL n. sp.
Holotype: USNM 1107794. A whole-mounted specimen, stained with hematoxylin; collected by S. Fend and D. Lenat.
Type locality: North Carolina: Richmond Co.: Drowning Creek at SR 1004, N35 º 03’ 58.3”, W79º 32’ 58.4”, 15 March 2007. Collected in fine gravel, in relatively fast current.
PARATYPES: USNM 1107795-1107801. All collected at the type locality, 15 March 2007 or 26 April 2007. 3 whole mounts, 2 dissected on slides, 2 sagittally sectioned.
Other material: North Carolina: Hoke Co.: Rockfish Creek at SR 1300, 1 May 1994. 4 dissected. Jones Co.: unnamed tributary to Holston Creek, 16 March 2007. 1 whole mount. Richmond Co.: from the type locality, 13 July 2001. 2 whole mounts. 15 March 2007. 4 dissected, 3 whole mounts, 2 sagittally sectioned. 26 April 2007. 8 dissected, 6 whole mounts. Bones Fork at SR 1487, 14 February 2007. 1 dissected. Swain Co.: Forney Creek in Great Smoky Mountains National Park, 28 April 2003. 1 whole mount. Virginia: Amherst Co.: Seep in Peaks of Otter area, 7 May 2007. 1 whole mount.
Etymology: The genus is named for Prof. Patrick Martin (Royal Belgian Institute of Natural Sciences), whose persistent efforts on behalf of obscure, intractable, deep-lake oligochaetes has provided much insight to lumbriculid systematics. The specific epithet refers to its general distribution within North Carolina.
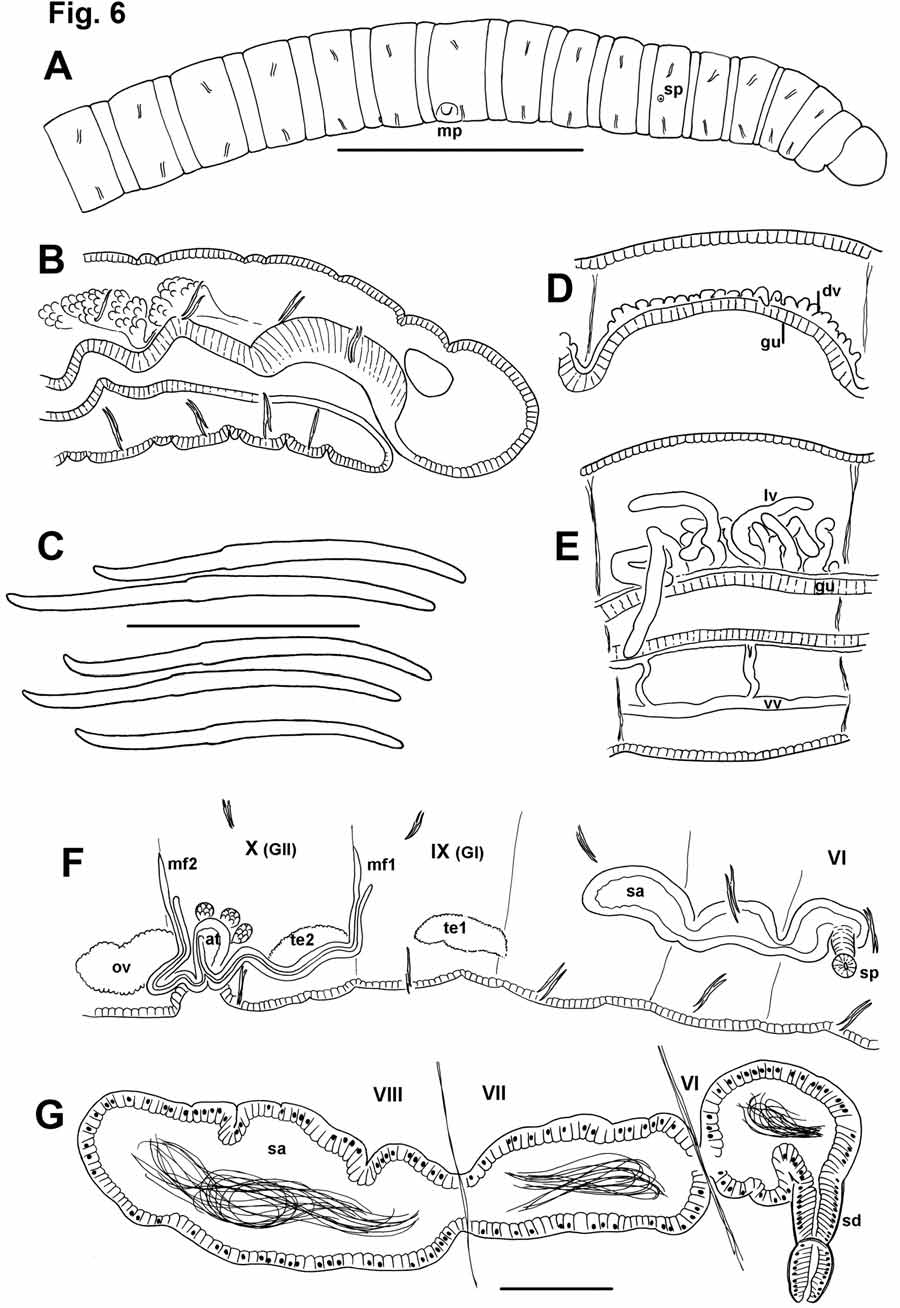
Description. Small worms, length of preserved specimens 11 to 24 mm (mean 18 mm); maximum diameter usually in clitellum, 0.36–0.67 mm (mean 0.48 mm); 48–80 segments (mean 66). Prostomium rounded, slightly shorter than wide, separated from the peristomium by a distinct groove ( Fig. 6 View FIGURE 6 A). Secondary segmentation consists of an anterior annulus in segments beginning with IV; in preclitellar segments the anterior annulus is 1/5 to 1/4 the segment length, but in postclitellar segments it is about 1/3 the segment length. Chaetae paired, in 4 bundles in each segment, beginning in II. Chaetae slightly sigmoid, simple-pointed, with nodulus slightly ectal to the midpoint, 0.32–0.43 the distance from the tip ( Fig. 6 View FIGURE 6 C). Chaetae in preclitellar segments near mid-segment, but in postclitellar segments they are near the posterior third. Chaetal length 70– 100 Μm in clitellar region, 55–65 Μm posteriorly.
Epidermis in prostomium 25–32 μm thick; in anterior segments 14–21 Μm; in clitellum 18–27 Μm. Clitellum usually from mid-IX to mid-XIII when fully developed, glandular, but only slightly thicker than normal epidermis. A distinct, dorsal pharyngeal pad of columnar cells in II to mid-III; when everted, only the dorsal pad is extruded (ventral part of pharynx is stationary). Dorsal gut wall is thickened behind the pharyngeal pad, but ventral pharynx wall is thin through mid-IV, to the beginning of the esophagus ( Fig. 6 View FIGURE 6 B). Pharyngeal extensor muscles join dorsal body wall at base of prostomium; more extensive retractors at dorsum in II–IV. Pharyngeal glands usually IV–VII.
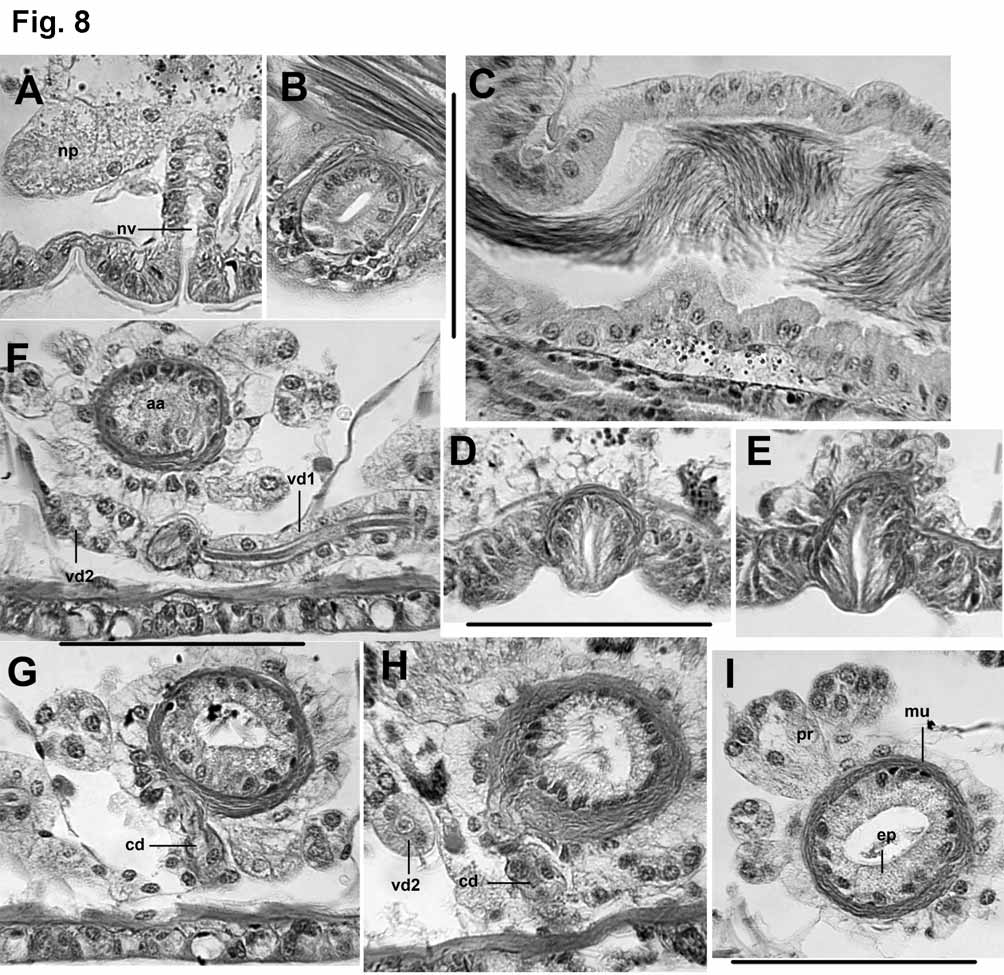
First pair of nephridia on 6/7. Nephridia absent in VIII–XII, usually paired in XIII, and occurring irregularly in posterior segments. Each nephridium with small anteseptal funnel, a granular postseptal expansion 50–120 μm long by 30–50 μm wide, and a convoluted duct that may pass through several anterior or posterior segments ventral to the gut, usually terminating in a narrow vesicle (up to 85 μm long by 24 μm wide) at the nephropore ( Fig. 8 View FIGURE 8 A).
Main dorsal and ventral blood vessels prominent, without secondary longitudinal vessels. Dorsal vessel separate from gut through about XV, connected to perivisceral sinus by short blood vessels; closely appressed to gut posterior to XV, eventually becoming indistinguishable from perivisceral sinus in most specimens. Circum-pharyngeal trunks join in V or at 4/5 to form the ventral vessel. One pair of commissural blood vessels in II–X; they are prominent and convoluted in II–VII, but shorter and difficult to see in IX–X. Perivisceral sinus begins in about VI; chloragogen begins in about VII. Starting near XXX, the dorsal vessel appears nodulate ( Fig. 6 View FIGURE 6 D); in posterior segments five or more very short, blind blood vessels appear dorsally on the dorsal vessel; by about XLV these unbranched, blind vessels are 1/4 to 1/2 as long as the body width ( Fig. 6 View FIGURE 6 E).
Male pores paired in X, behind chaetae on ventral chaetal lines, usually conspicuous on short (10–15 μm) papillae within shallow pits 50–70 μm wide; the pits formed by a ring of non-glandular epidermal cells ( Fig. 8 View FIGURE 8 D–E). Female pores small, paired, on chaetal line at intersegmental groove 11/12. Spermathecal pores usually conspicuous on lateral lines near midpoint of VI, as a small slit within ectal end of the duct, diameter 35 μm ( Fig. 8 View FIGURE 8 B). Paired testes on anterior septa in IX and X. Ovaries paired in XI. Sperm sacs rarely extend anteriorly into VIII; posteriorly they extend as far as XIV–XVI. Egg sacs extend as far as XVI–XVIII.
Spermathecal ducts 85–140 μm long ( Fig. 6 View FIGURE 6 G). Main (ental) section about 50–90 μm long by 50–60 μm wide at ental end, with thick epithelium, tapering to a narrower neck; a short ectal section has more cuboidal epithelial cells, and a distinct muscle layer (up to 2 μm thick). Spermathecal ampulla long and sacciform, constricted at septa, usually extending to VIII ( Fig. 6 View FIGURE 6 F–G). Lining cells not vacuolated or differentiated in different parts of the ampulla; all cuboidal, thickness variable but relatively uniform within an individual, 5–25 μm. Length of ampulla about 500–1000 μm, maximum diameter to 100–250 μm. Sperm loosely packed and uniformly distributed throughout ampulla ( Fig. 6 View FIGURE 6 G, 8C).
Male funnels paired on 9/10 and 10/11; anterior lips bent forward; posterior lips appressed to septum. Anterior funnel 60–90 μm high; posterior 75–120 μm. Anterior vas deferens averages about 500 μm long; posterior slightly shorter, averaging about 400 μm, forming a posterior loop extending as far as mid-XI ( Fig. View FIGURE 7
7A). Both vasa deferentia up to 20–28 μm thick, with ciliated lumen about 5–10 μm wide; vasa narrow to 16– 24 μm as they join a short common duct. The common duct is 50–90 μm long and usually thinner than the vasa deferentia (16–22 μm); it has a thin muscle coat (1–2 μm), a narrow lumen (3 μm), usually without cilia, and joins the atrium apically or subapically, always from the posterior-median side ( Fig. 7 View FIGURE 7 A–D, Fig. 8 View FIGURE 8 G–H).
Atria with a pyriform ampulla 60–120 μm long by 55–85 μm wide, narrowing to a weakly differentiated ectal duct about 70 μm long by 30–40 μm wide ( Fig. 7 View FIGURE 7 C). Atrial ampulla with an outer muscle layer 5–12 μm thick, an inner epithelium 12–20 μm thick, and a narrow lumen to 10–25 μm wide ( Figs. 8 View FIGURE 8 G–I). Ampulla loosely covered by a layer of non-granular cells and a variable number (less than 10 to more than 25) of petiolate prostate glands 30–70 μm long ( Fig. 8 View FIGURE 8 I). Prostate cells usually very granular and often indistinct; up to 30 cells in the largest bundles, but usually fewer. Transition between the ampulla and duct is gradual; duct has thinner muscle layer (2 μm), and lacks prostates. Ectally, the duct terminates in a slightly thickened penial structure 30–50 μm high. The penial structure has somewhat elongated cells, forming a distinct papilla in most specimens ( Figs. 7 View FIGURE 7 C; 8D–E).
Remarks. The unusual common duct joining the vasa deferentia to the atrium is thin and difficult to observe. In most cases it is histologically distinct from the vasa deferentia in its narrow and irregular form, the apparent lack of cilia, and the thin muscle coat. However, these characters all vary, and cilia appear to be present in at least one specimen.
The single, geographically isolated specimen from Peaks of Otter, Virginia, is somewhat distinctive. There appears to be no papilla at the male pore, and the common duct joins the atrium well before the apex ( Fig. 7 View FIGURE 7 D). Although the Forney Creek population also appears disjunct, the single specimen appears quite similar to the type material.
The placement of spermathecal pores ( Figs. 6 View FIGURE 6 A, 9A) makes mature individuals of both Martinidrilus species easy to identify as whole-mounted specimens. The many short, blind vessels in posterior segments ( Figs. 6 View FIGURE 6 E, 9B) distinguish Martinidrilus from all confirmed Nearctic lumbriculids; therefore, cleared, immature specimens can also be identified to genus. Although Trichodrilus allobrogum Claparède also may have multiple lateral blood vessels (see below), the only North American record attributed to that species is doubtful, and blood vessels could not be described in that specimen ( Kindred 1918; Cook 1971a).

| USNM |
Smithsonian Institution, National Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |