
Macrodiplosis selenis Kim & Yukawa
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3821.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:E68F3711-DB8E-4D6D-807A-BEA6275DC1B8 |
|
DOI |
https://doi.org/10.5281/zenodo.5629583 |
|
persistent identifier |
https://treatment.plazi.org/id/03DBD509-FFB2-354F-CFC4-DD7EFAF8D61E |
|
treatment provided by |
Plazi |
|
scientific name |
Macrodiplosis selenis Kim & Yukawa |
| status |
sp. nov. |
Macrodiplosis selenis Kim & Yukawa View in CoL sp. nov.
Figs 9–21 View FIGURES 9 – 13 View FIGURES 14 – 21 .
MALE ( Figs 9–10 View FIGURES 9 – 13 ) Eye bridge about 6 facets long medially. Antenna: scape with 1 to 4 setae (n=29); pedicel with 4 to 7 setae (n=4); basal enlargement of flagellomere V about 0.87 times as long as wide; distal enlargement 1.3–1.9 times as long as wide, distal neck about 1.8 times as long as intermediate neck ( Fig. 9 View FIGURES 9 – 13 ).
Wing length: 2.1–2.5 mm, 2.6–3.1 times as long as wide. Legs densely covered with brown hairs. Tarsal claw simple on all legs, as long as empodium. See Table 4 View TABLE 4 for detailed data on setal counts and measurements.
Male Female ......continued on the next page Male Female Abdominal tergites I to VI rectangular, wider than long, with hair-like scales, a pair of trichoid sensillae, one to two caudal rows of setae and many lateral setae at midlength; tergite VII rectangular, narrower than tergite VI, with a pair of trichoid sensillae and many scattered setae mediolaterally; tergite VIII unpigmented, bare, with a pair of trichoid sensillae. Abdominal sternites II to VI rectangular, wider than long, without any scales, with a pair of trichoid sensilla, one to two caudal rows of setae and many scattered setae medially; sternite VII rectangular, with a pair of trichoid sensillae and many scattered setae mediolaterally; sternite VIII rectangular, with a pair of trichoid sensillae and a few scattered setae medially.
Terminalia as in figure 10; cerci short, with many short setae all over and some longer setae distally; hypoproct as long as cerci, much shorter than aedeagus, distally concave; lateral lobes directed ventro-distally, with several longer setae distally; gonocoxite cylindrical, without mediobasal lobe; gonostylus relatively massive basally, distally attenuate, with many short and some longer setae, setulose except on attenuate portion, with comblike apical tooth; aedeagus tapered to apex.
FEMALE ( Figs 11–13 View FIGURES 9 – 13 ). Eye bridge 6 to 7 facets long medially ( Fig. 11 View FIGURES 9 – 13 ). Basal enlargement of flagellomere V about 2.2 times as long as wide ( Fig. 12 View FIGURES 9 – 13 ).
Wing length: 2.6–2.8 mm, 2.5–3.4 times as long as wide. See Table 4 View TABLE 4 for detailed data on setal counts and measurements.
Abdominal tergites I to VI as in male; tergite VII rectangular, wider than long, vestiture similarly arranged; tergite VIII unpigmented, bare. Abdominal sternites II to VI as in male; sternite VII rectangular, wider than long, with many scattered setae mediolaterally; sternite VIII undifferentiated. Ovipositor ( Fig. 13 View FIGURES 9 – 13 ) soft and pliable; cerci not fused, apically rounded.
MATURE LARVA ( Figs 14–20 View FIGURES 14 – 21 ). Reddish orange in body color. Antennal segment II about 22.3 µm long (n=3); cervical papillae without seta.
Thorax: spiracle present on prothorax; 6 dorsal papillae present on all thoracic segments ( Fig. 14 View FIGURES 14 – 21 ); 2 pleural papillae present on each side of all thoracic segments; all dorsal and pleural papillae with tiny setae. Sternal spatula on ventral surface of prothorax 149.8 to 212.8 µm (n=3), distally rather deeply incised by U shaped emargination, forming pair of subglobular lobes; shaft of sternal spatula proximally attenuated ( Figs 15–16 View FIGURES 14 – 21 ); 6 lateral papillae in 2 clusters of 3 each, present on each side of midline, 2 papillae in each cluster with a minute seta, asetose papillae in each cluster obscure, not clearly present in the photograph, but visible under a phase-contrast microscope with high magnification (common in many cecidomyiid larvae; see Fig. 7 View FIGURES 5 – 8 in Möhn (1955) for M. dryobia ) ( Fig. 17 View FIGURES 14 – 21 ); 2 sternal papillae present on each thoracic segment, without setae; 2 inner pleural papillae on all thoracic segments without setae.
Abdomen: 6 dorsal papillae present on abdominal segments I–VII ( Fig. 18 View FIGURES 14 – 21 ); 2 pleural papillae present on each side of abdominal segments I–VII; all dorsal and pleural papillae with tiny setae; 2 dorsal papillae present on abdominal segment VIII, with minute setae, 2 pleural papillae present on abdominal segment VIII; 2 anterior ventral papillae and 2 posterior ventral papillae without setae on abdominal segments I–VII; 2 ventral papillae of abdominal segment VIII without setae; 4 of 8 terminal papillae setose, with setae 17.3 to 30.8 µm long, remaining 4 corniform ( Fig. 19 View FIGURES 14 – 21 ); anal papillae not apparent ( Fig. 20 View FIGURES 14 – 21 ).
PUPA ( Fig. 21 View FIGURES 14 – 21 ). Base of antenna short; cephalic pair of setae long, 88.4 to 105.9 µm in length (n=2); frons smooth; lower facial papillae absent; 3 pairs of lateral facial papillae present, one pair with tiny setae; prothoracic spiracle long, 196.7 to 211.7 µm in length (n=3).
Abdominal spiracles short, present on abdominal segments II–VI; abdominal segments II–VIII with several macrospines on central area of anterior third of dorsal surface; other parts of dorsal surface on abdominal segments II–VIII covered with microspines; a few dorsal papillae visible on abdominal segments II–VIII without setae; pleural papillae not apparent.
Material examined. Holotype: ♂ (on slide, Cecid. No. KW001) emerged on 13 April 2012 from a gall on Q. serrata collected by W. Kim on 18 October 2011 from Mitsusawa, Ogori City, Fukuoka Prefecture, Japan. Paratypes: 3 ♂♂, 3 ♀♀ & 3 pupae (on slides, Cecid. No. KW002-010), same data as holotype; 4 mature larvae (on slides, Cecid. No. KW011-014) taken out of galls on Q. serrata collected on 6 October 2011 from Mitsusawa by W. Kim.
Distribution. [ Japan] Hokkaido, Honshu (Ibaraki, Nagano and Hyogo Prefectures), Shikoku (Kagawa Prefecture) and Kyushu (Fukuoka, Oita and Miyazaki Prefectures), and [ Korea] Gyeongsangbukdo.
Host plants. Quercus serrata , Q. dentata and Q. mongolica (Fagaceae) .
Gall shape. The leaf margin between two successive serrations is folded upward by the larvae of M. selenis to form a crescent moon-shaped gall (Figs 1–3). This gall is similar in appearance to that induced by M. roboris on Quercus species in Europe ( Fig. 5 View FIGURES 5 – 8 ). These galls differ from those folded downward by M. pustularis on Quercus spp. in Europe and Macrodiplosis sp. 2 on Q. mongolica in Japan and South Korea ( Figs 6–7 View FIGURES 5 – 8 ).
Etymology. The specific name, selenis , is derived from the Greek for “crescent moon”, which resembles the shape of the upwardly folded leaf-margin.
Remarks. Macrodiplosis selenis is morphologically distinguished from M. roboris in having a distinctly longer distal neck of male flagellomere V ( Fig. 9 View FIGURES 9 – 13 versus Fig. 569 in Skuhravá 1997). M. selenis differs from M. pustularis in having a narrower aedeagus ( Fig. 10 View FIGURES 9 – 13 versus Fig. 23 View FIGURES 22 – 25 ) and a constricted basal enlargement of female flagellomere V ( Fig. 12 View FIGURES 9 – 13 versus Fig. 24 View FIGURES 22 – 25 ). M. selenis cannot be distinguished from M. pustularis by other characteristics of the adult ( Figs 22, 25 View FIGURES 22 – 25 ) and pupa ( Figs 27–28 View FIGURES 26 – 28 ). The proximal part of the larval sternal spatula is distinctly widened in M. roboris ( Fig. 29 View FIGURES 29 – 33 ) and slightly widened in M. pustularis ( Fig. 26 View FIGURES 26 – 28 ), while it is attenuated into a simple end in M. selenis ( Fig. 15 View FIGURES 14 – 21 ), which was consistent also in all 20 full-grown larvae of M. selenis examined before genetic analysis (Table 3). Other larval characteristics of M. selenis ( Figs 17, 19, 20 View FIGURES 14 – 21 ) were similar to those of M. roboris ( Figs 29–33 View FIGURES 29 – 33 ). M. selenis differs from the Russian Far East species, M. flexa , in having the following features: lobes of cerci triangular; distal margin of hypoproct bilobed with widely U-shaped emargination; and aedeagus distally attenuated ( Fig. 10 View FIGURES 9 – 13 versus Fig. 18 View FIGURES 14 – 21 in Kovalev 1972). Other adult characteristics of M. selenis ( Figs 9, 11–13 View FIGURES 9 – 13 ) are similar to those of M. flexa ( Figs 8–17 View FIGURES 5 – 8 View FIGURES 9 – 13 View FIGURES 14 – 21 in Kovalev 1972). Two other Palaearctic congeners, M. inflexa ( Bremi) (Bremi 1847) and M. quercus ( Loew) (Loew 1850) were insufficiently described for us to compare with M. selenis . The type of the former may be lost and the name of the latter has not been used since originally named, except for a reference in Löw (1877). Macrodiplosis quercus is considered to be nomina dubia (as Cecidomyia quercus ; Gagné & Jaschhof 2014).
Genetic analysis. The strict consensus tree of 197 equally parsimonious trees (tree length 423, consistency index = 0.6, retention index = 0.90, rescaled consistency index = 0.54) clearly indicated that the Palaearctic species, M. pustularis , M. roboris , M. selenis and Macrodiplosis sp. 2, were each included in separate clades ( Fig. 34 View FIGURE 34 ). Unidentified specimens collected from Poland proved to be identical to M. pustularis . Each clade was supported by a 100% bootstrap value. In contrast, Nearctic species could not be identified to the species level because of confusion in old descriptions (Gagné 2012, personal communication), and they were not placed in a single clade, although the strict consensus tree reflected some of the differences in host plant species ( Fig. 34 View FIGURE 34 ).
Five haplotypes were recognized among individuals of M. selenis . Estimated mean sequence divergences between M. selenis and other Palaearctic species are as follows: M. roboris , 42 bp (6.5%); M. pustularis 50 bp (7.6%); Macrodiplosis sp. 2 on Q. dentata and Q. mongolica in Hokkaido, Japan 57 bp (8.7%). The mean sequence divergence was 85 bp (12.9%) between the Nearctic and Palaearctic species.
Life history. The dissection of galls revealed that larvae of M. selenis remained in the galls throughout the summer and dropped to the ground in October and November, where they overwintered. Thus, M. selenis is univoltine, as has been noted for M. pustularis and M. roboris (e.g., Skuhravá et al. 2006; David 2012). The detailed life history strategy and other ecological traits of M. selenis will be published in a separate paper.
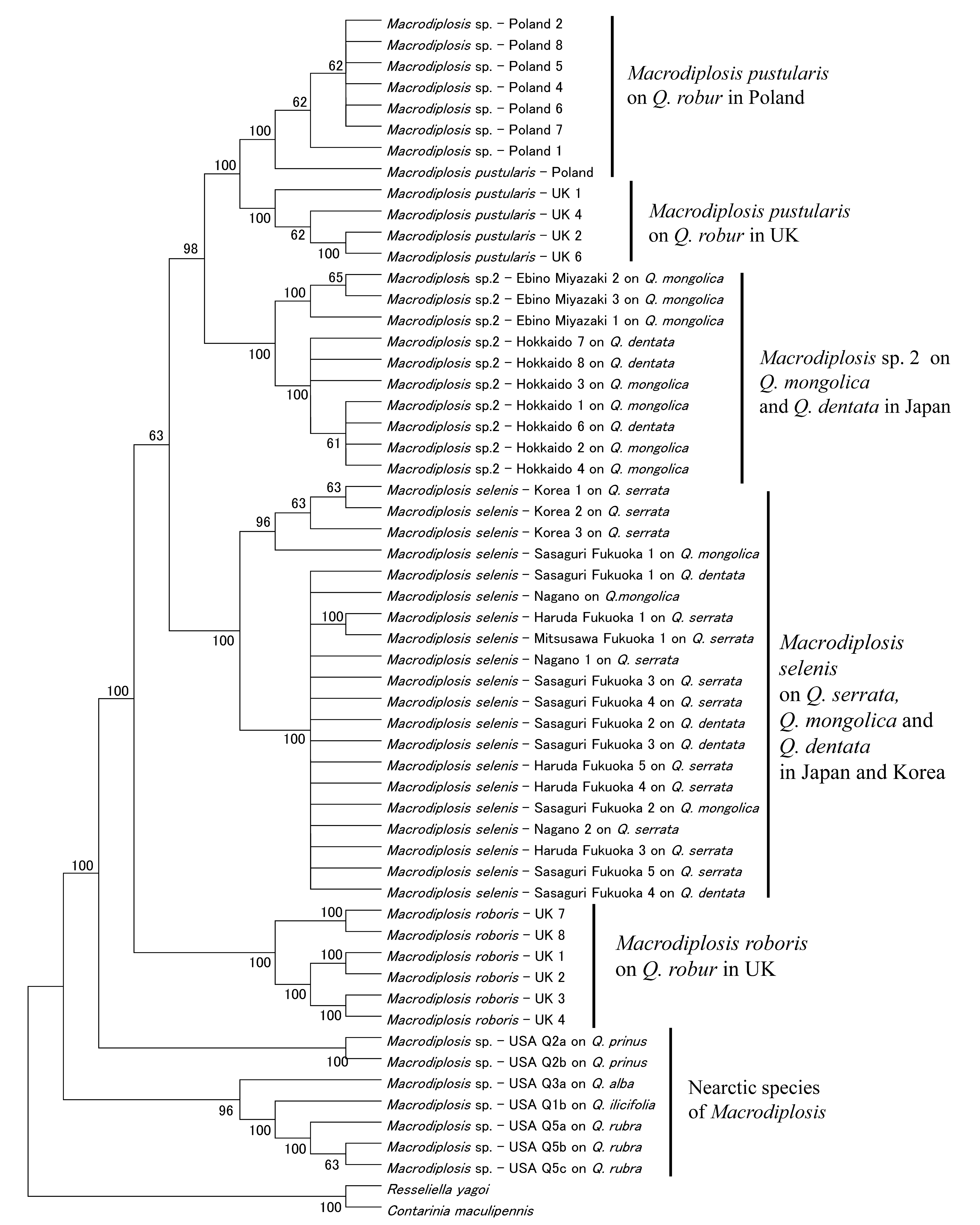
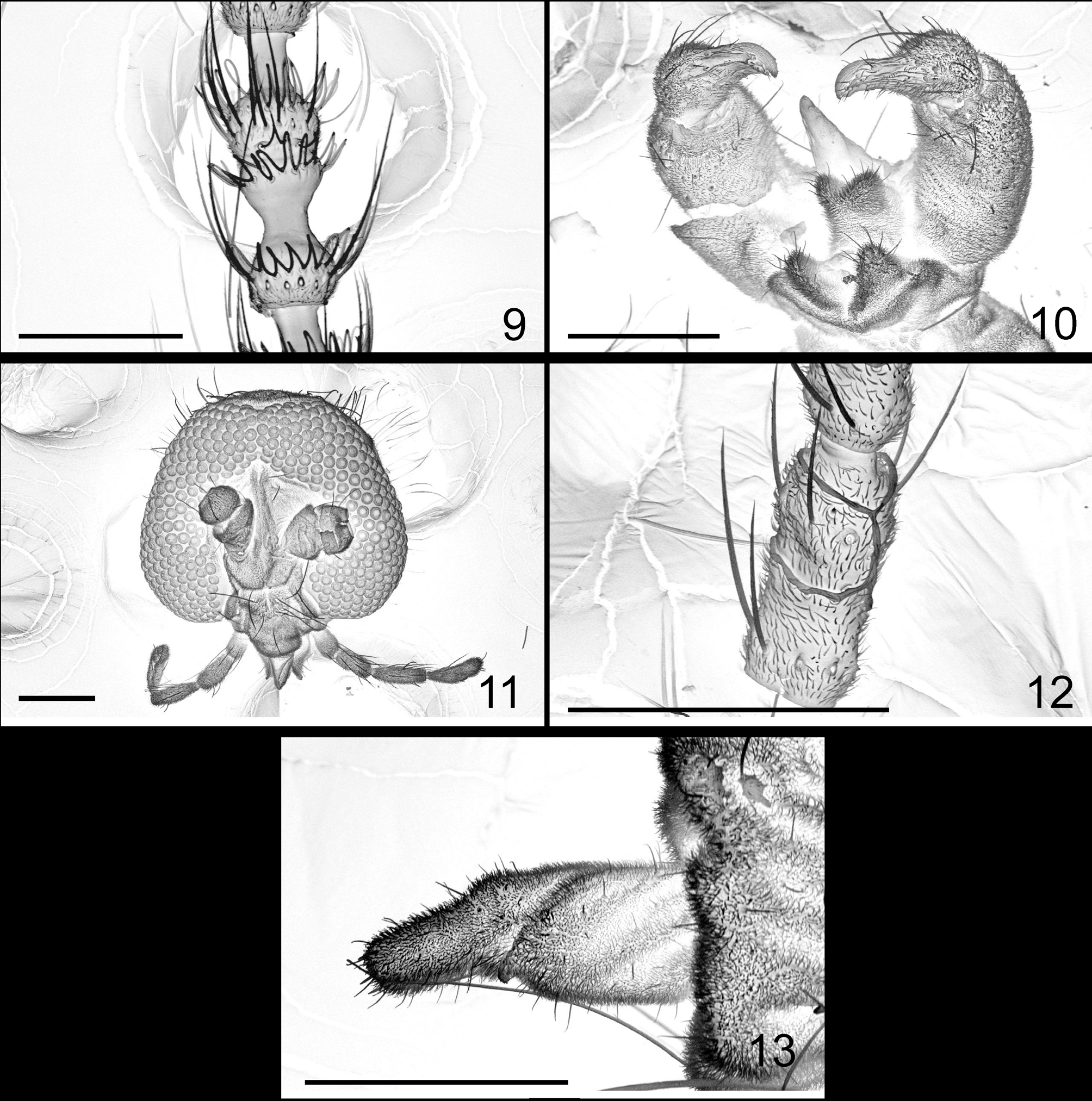


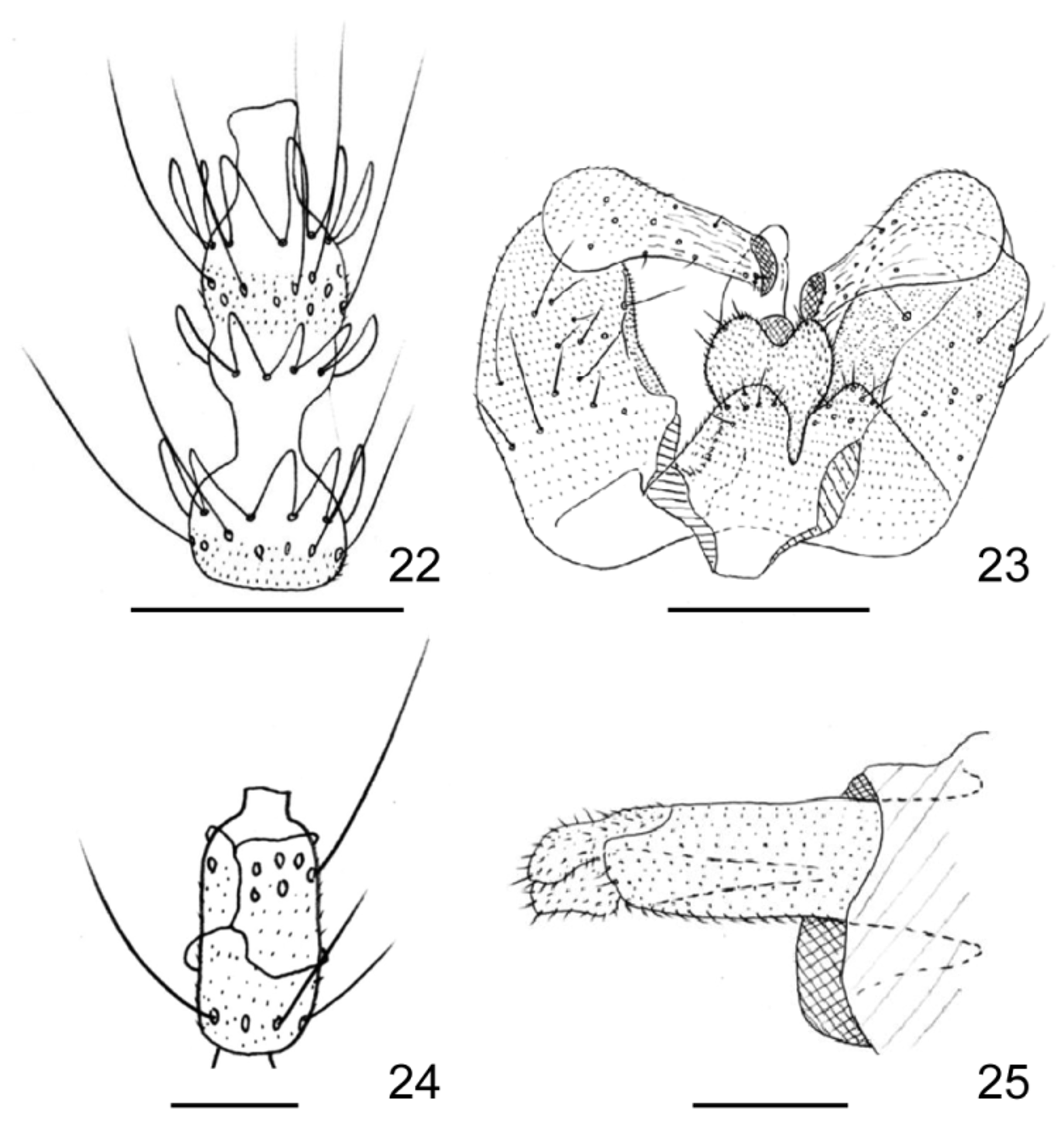
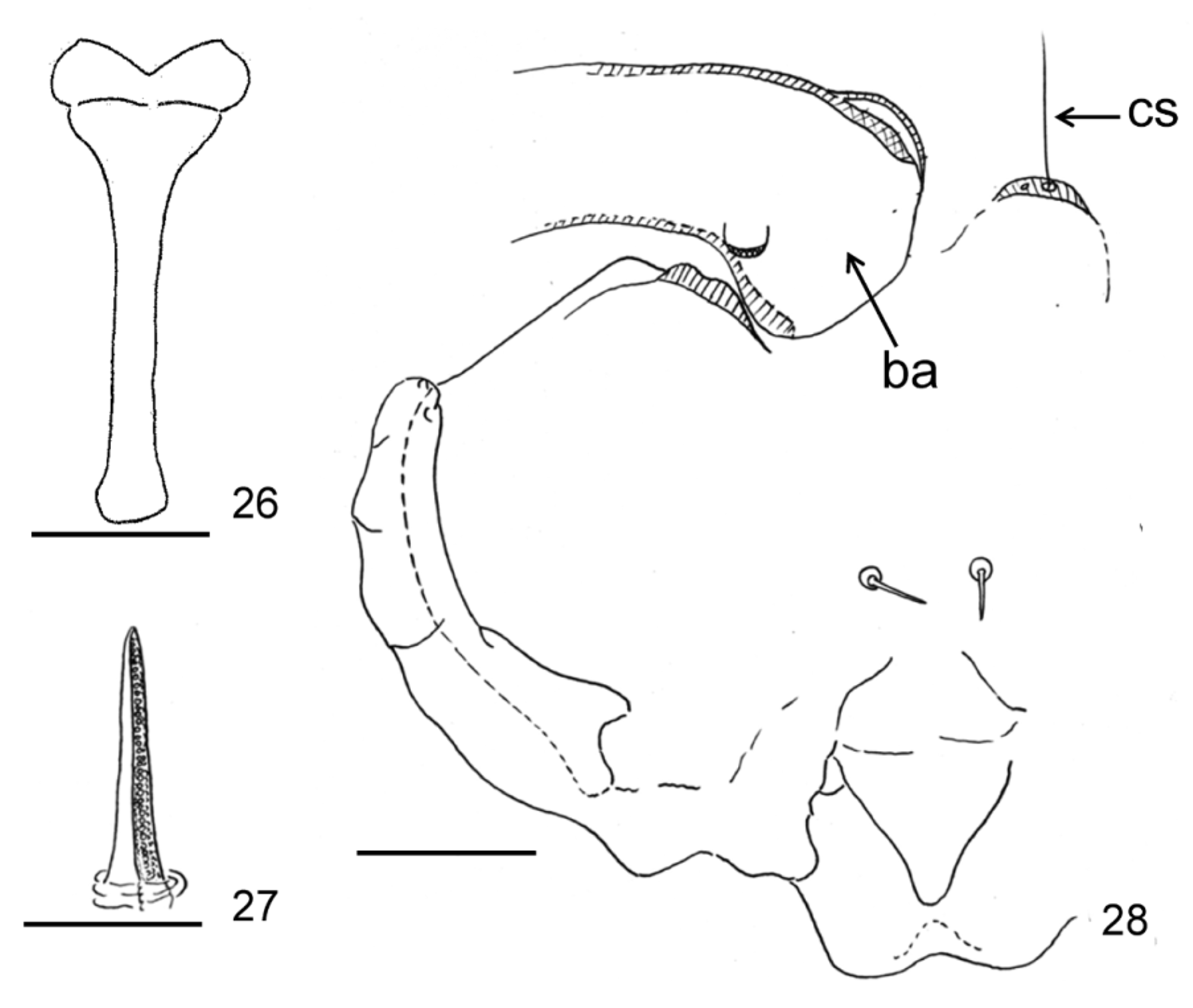
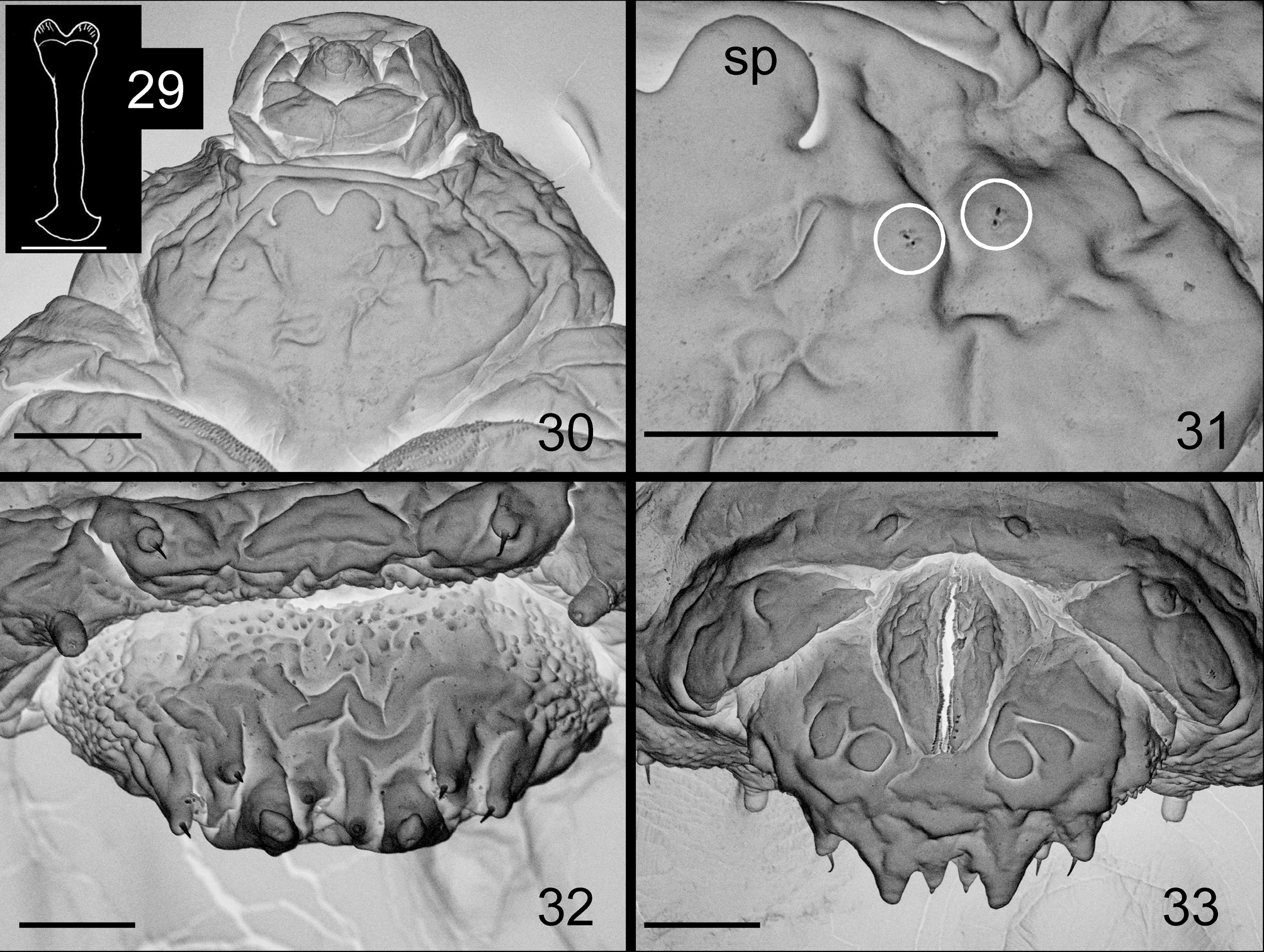
TABLE 4. Frontoclypeal and mesepimeral setal counts, and measurements of palpus (µm), flagellomere III (µm), wing (mm) and legs (mm) of Macrodiplosis selenis sp. n.
| n | Mean±SD | (range) | N | Mean±SD | (range) | |
|---|---|---|---|---|---|---|
| Frontoclypeal setae | 4 | 8.0 ± 0.8 | (7.0–10.0) | 3 | 8.3 ± 2.9 | (5.0–10.0) |
| Mesepimeral setae | 4 | 7.8 ± 2.2 | (5–10) | 3 | 9.3 ± 0.6 | (9–10) |
| Palpus Segment I | 4 | 53.4 ± 7.3 | (45.8–62.5) | 3 | 50.3±7.5 | (42.5–57.5) |
| Segment II | 4 | 61.8 ± 7.0 | (52.0–68.7) | 3 | 62.8±6.8 | (55.7–69.2) |
| Segment III | 4 | 69.3 ± 18.3 | (51.8–89.2) | 3 | 76.7±10.3 | (65.5–85.9) |
| Segment IV | 3 | 91.3 ± 25.3 | (63.3–112.5) | 3 | 116.6±30.6 | (83.7–144.2) |
| Flagellomere III | ||||||
| Length of basal enlargement | 4 | 41.6 ± 2.6 | (39.7–45.2) | 3 | 112.7±3.1 | (109.4–115.4) |
| Width of basal enlargement | 4 | 61.9 ± 2.8 | (59.2–65.8) | 3 | 58.8±4.4 | (55.7–63.8) |
| Length of intermediate neck | 4 | 28.1 ± 2.5 | (25.0–30.8) | - | - | - |
| Length of distal enlargement | 4 | 65.3 ± 2.6 | (63.3–69.2) | - | - | - |
| Width of distal enlargement | 4 | 61.2 ± 2.6 | (57.5–63.7) | - | - | - |
| Length of distal neck | 4 | 39.2 ± 3.9 | (35.0–44.2) | 3 | 17.1±1.1 | (15.8–18.0) |
| Wing length | 4 | 2.3 ± 0.2 | (2.1–2.5) | 3 | 2.7 ± 0.1 | (2.6–2.8) |
| Wing width | 4 | 0.9 | (0.9–1.0) | 3 | 1.1 | (1.0–1.1) |
| Length/width | 4 | 2.5 ± 0.1 | (2.4–2.6) | 3 | 2.6 ± 0.1 | (2.4–2.7) |
| Fore leg Femur | 3 | 1.0 | (0.9–1.0) | 3 | 0.9 ± 0.1 | (0.9–1.0) |
| Tibia | 3 | 0.9 ± 0.1 | (0.8–1.0) | 3 | 1.0 ± 0.1 | (0.9–1.0) |
| Tarsomere II | 3 | 0.8 | (0.8–0.9) | 3 | 0.8 | (0.8–0.8) |
| Tarsomere III | 3 | 0.4 | (0.4–0.4) | 3 | 0.4 | (0.4–0.4) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |