
Kinnecaris esbe, Karanovic & Cooper, 2011
|
publication ID |
https://doi.org/10.11646/zootaxa.3026.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.5285523 |
|
persistent identifier |
https://treatment.plazi.org/id/03924C3A-FFB2-A60C-FF41-FD7CA985FB1E |
|
treatment provided by |
Felipe |
|
scientific name |
Kinnecaris esbe |
| status |
sp. nov. |
Kinnecaris esbe sp. nov.
( Figs. 7–10 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
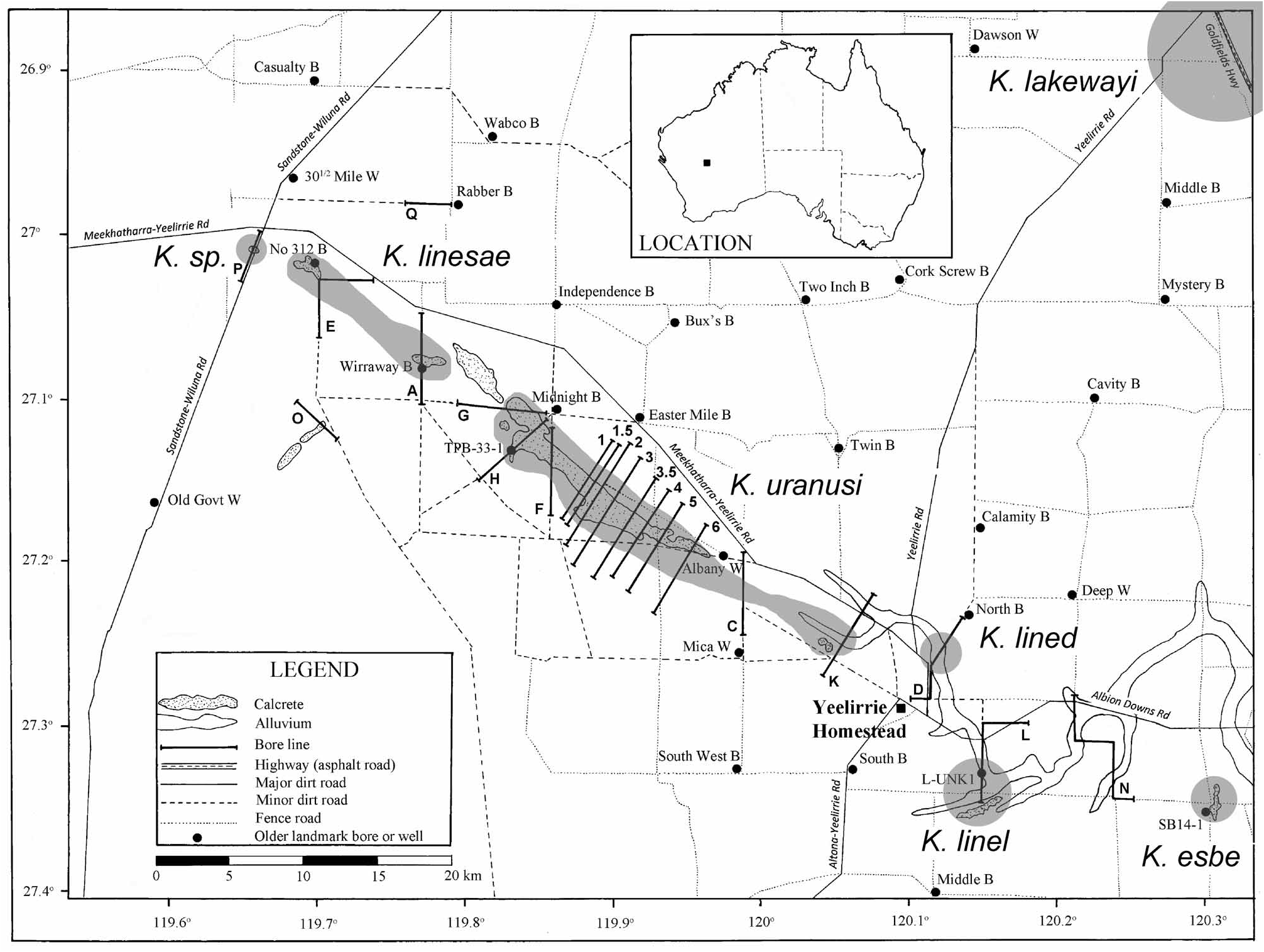
Type locality. Australia, Western Australia, Yilgarn region, Yeelirrie pastoral station, bore SB14-1, 27.344283˚ S 120.307708 ˚E (south-eastern corner on Fig. 24 View FIGURE 24 ) .
Type material. Holotype male, allotype female, and one paratype female on one SEM stub in toto coated with carbon ( WAM C47185); one paratype male dissected on one slide ( WAM C47186); one paratype female dissected on one slide ( WAM C47187); one paratype female destroyed for DNA sequence (amplification unsuccessful); 13 paratypes ( one male + seven females + five copepodids) in alcohol; all collected at type locality, leg. T. Karanovic & S. Callan, 18 March 2010, seLN8180. Other paratypes: One female destroyed for DNA sequence (amplification unsuccessful); one male + three females + five copepodids in alcohol; all collected at type locality, leg. T. Karanovic & G. Perina, 16 March 2010, seLN8517.
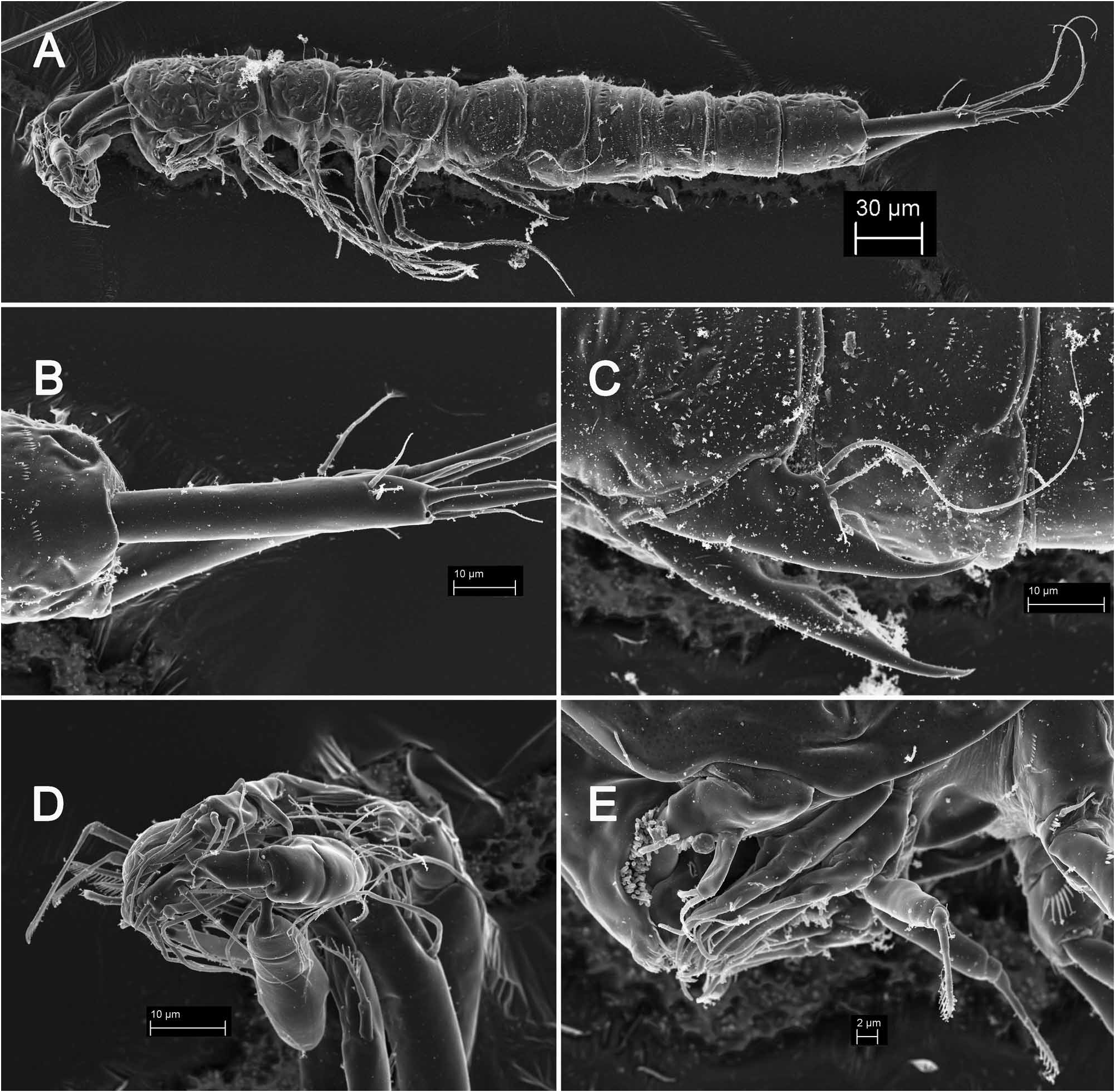
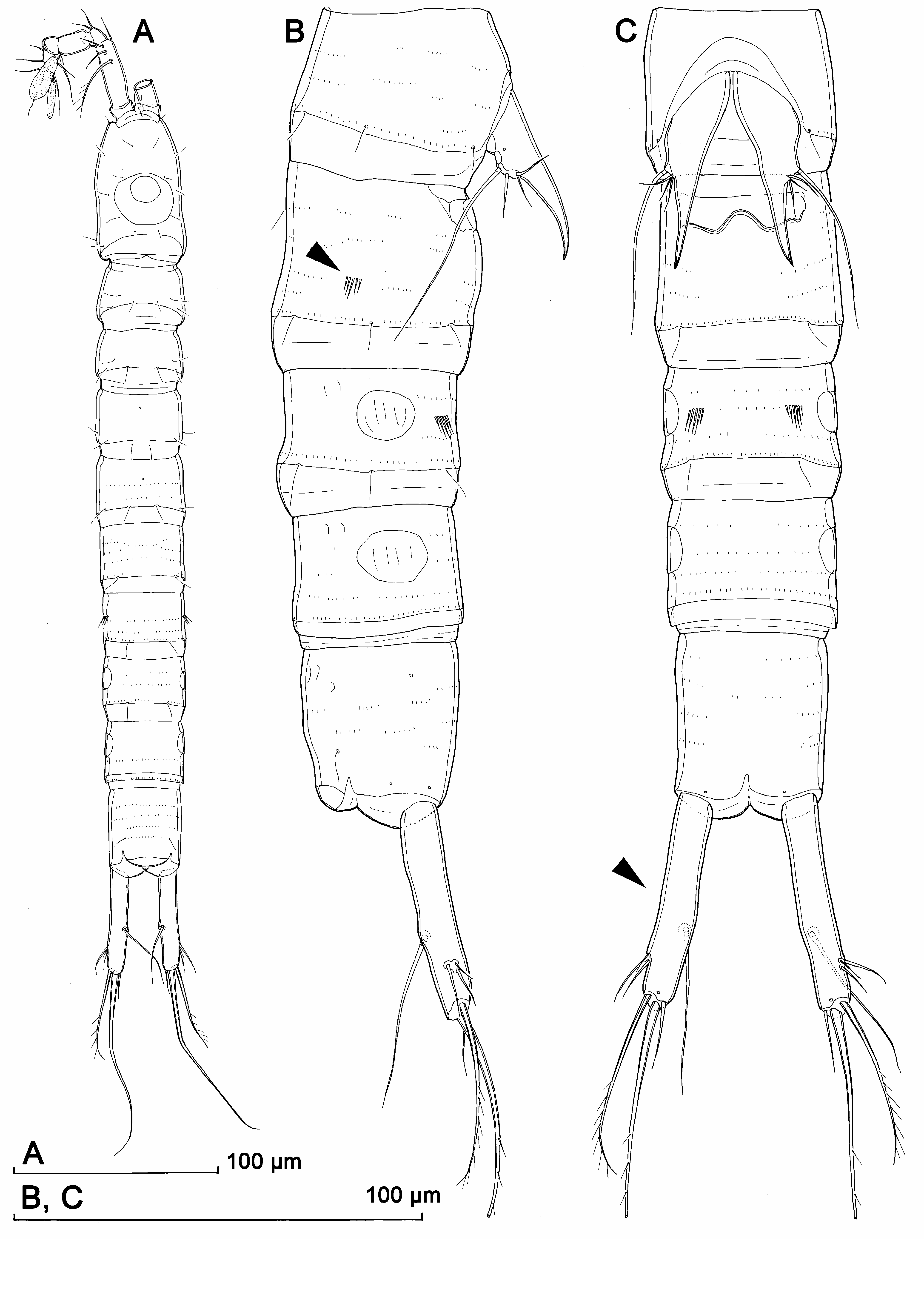
Description. Male (based on holotype and several paratypes). Total body length from 374 to 425 µm (374 µm in holotype). Surface of integument of all somites with relatively dense and shallow cuticular pits, and all somites after cephalothorax with numerous short rows of minute spinules; third and fourth urosomites with some additional larger spinules ventrally and/or laterally ( Figs. 7A, C View FIGURE 7 , 8A View FIGURE 8 ). Colour, nauplius eye, rostrum, body segmentation, and pore and sensilla pattern of all somites as in Kinnecaris lakewayi (see above). Habitus ( Fig. 7A View FIGURE 7 ) cylindrical and very slender, without any demarcation between prosome and urosome dorsally, but with urosome wider in lateral view; prosome/urosome ratio about 0.6; greatest width from dorsal view at posterior end of cephalothorax, but very hard to establish. Body length/width ratio about 8.9; cephalothorax only slightly wider than genital somite in dorsal view. Integument more strongly sclerotized than in previous two species, and with relatively deep and irregular depressions on all somites but especially on cephalothoracic shield and pleuras and tergites of three free prosomites ( Fig. 7A, B View FIGURE 7 ). Cephalothorax with clearly visible ( Fig. 7A View FIGURE 7 ) double dorsal cuticular window posteriorly; fourth and fifth urosomites each with pair of lateral circular windows, both clearly visible and that on fifth urosomite slightly larger ( Figs. 7A View FIGURE 7 , 8A View FIGURE 8 ).
Cephalothorax ( Fig. 7A View FIGURE 7 ) about 1.5 times as long as wide in dorsal view; representing about 17% of total body length. Surface of cephalic shield, tergites and pleuras of free pedigerous somites ornamented as in K. lakewayi , except for presence of sparse and shallow cuticular pits and many more minute spinules on free pedigerous somites ( Fig. 7A, C View FIGURE 7 ); many of short rows of minute spinules arched, especially on lateral sides ( Fig. 7C View FIGURE 7 ), while those on ventral side of urosomites tend to be straight, parallel, and sometimes join into longer rows ( Fig. 8A View FIGURE 8 ).
Genital somite ( Fig. 7A, C View FIGURE 7 ) ornamented with more than forty dorso-lateral short rows of spinules (each from three to nine spinules), in addition to three pairs of sensilla on posterior margin (one dorsal, one dorso-lateral, and one ventro-lateral), and one ventro-lateral pair of very small cuticular pores in anterior part (i.e., pore and sensilla pattern as in K. lakewayi ); no large spinules on this somite; spermatophore visible inside and similar in size to that in K. lakewayi .
Third urosomite ( Fig. 7A View FIGURE 7 ) with four rows of large spinules in anterior half (two ventro-laterally and two dorsolaterally; each with five to six spinules and central ones largest), in addition to six posterior sensilla, and many rows of minute spinules.
Fourth urosomite ( Fig. 7A View FIGURE 7 ) ornamented with two or three ventro-lateral large spinules ventrally of large and slightly swollen lateral cuticular window; additional ornamentation represented by six posterior sensilla and numerous minute spinules.
Fifth (preanal) urosomite ( Figs. 7A View FIGURE 7 , 8A View FIGURE 8 ) with larger and more more swollen windows but without any large spinules, sensilla or pores; only ornamentation sparse and shallow cuticular pits and numerous minute spinules, forming nearly continuous row along hyaline fringe.
Anal somite ( Figs. 7A, B View FIGURE 7 , 8A View FIGURE 8 ) ornamented with pair of large dorsal sensilla at base of anal operculum, pair of lateral large cuticular pores (one pore on each side) in anterior half, two pairs of minute cuticular pores laterally close to posterior margin, and pair of slightly larger ventral cuticular pores at base of caudal rami, in addition to hardly visible cuticular pits and many rows of minute spinules. Anal operculum less developed than in K. lakewayi and bent slightly towards ventral side, unornamented on outer surface, with row of slender spinules on inner surface, convex and smooth distal margin, almost reaching posterior end of anal somite, representing 68% of somite width. Anal sinus less widely opened than in K. lakewayi , with two diagonal rows of slender spinules on ventral side and transverse row of spinules on dorsal side (below anal operculum).
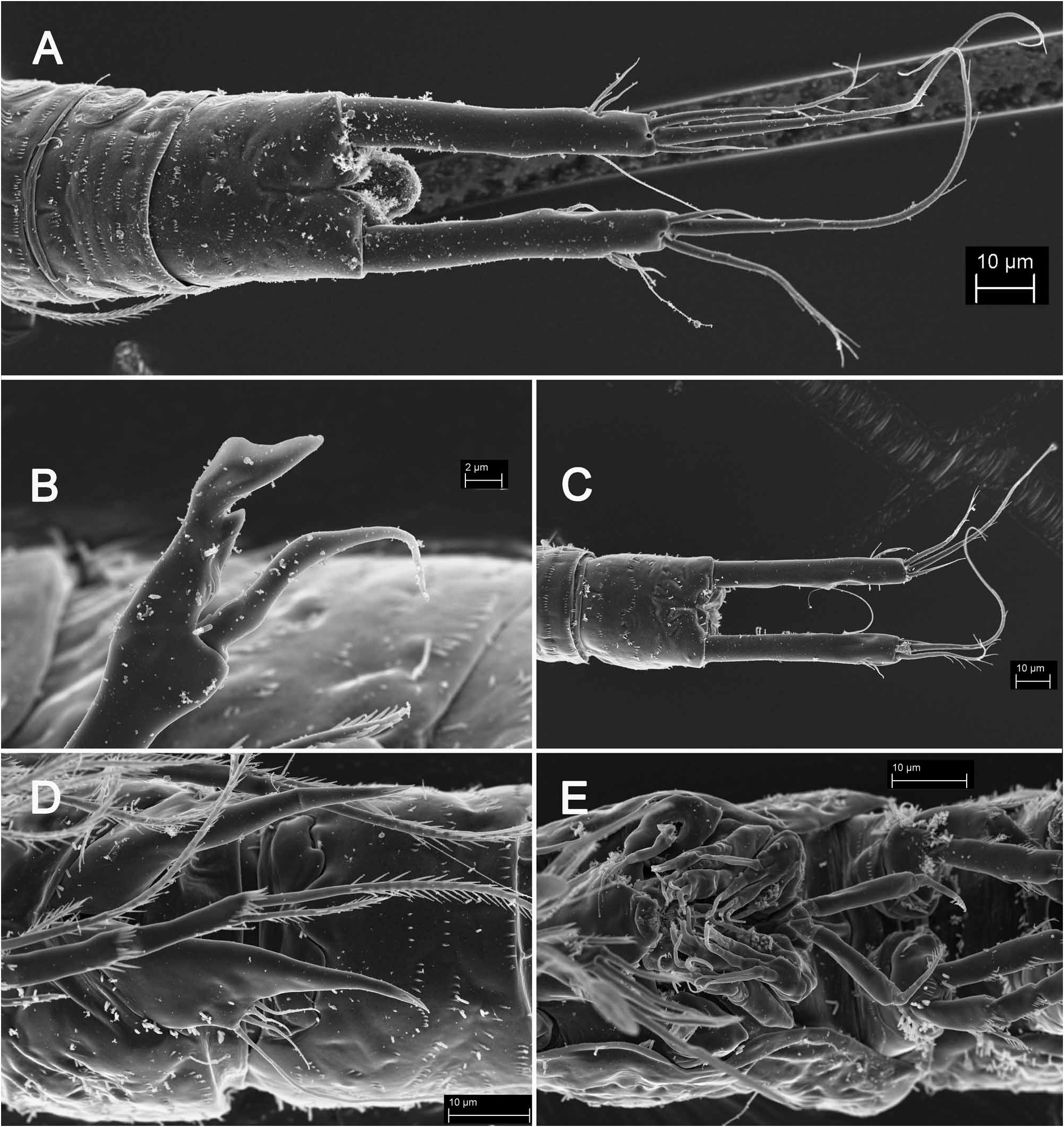
Caudal rami ( Figs. 7B View FIGURE 7 , 8A View FIGURE 8 ) almost 6.3 times as long as greatest width (dorsal view) and 1.3 times as long as anal somite, cylindrical in all views, nearly parallel, with space between them about 1.5 times one ramus’ width. Armature and ornamentation as in K. lakewayi , but due to the extreme elongation of the rami, the position of dorsal and lateral setae and their relative lengths differs. Dorsal seta slender and smooth, inserted slightly closer to inner margin at about 2/3 of ramus length, only 0.7 times as long as caudal ramus, triarticulate at base. Lateral setae thin and smooth, inserted close to each other at 5/6 of ramus’ length, in one depression with two positioned proximally and one distally; proximal seta which inserted more dorsally strongest and longest of three, but only about 0.2 times as long as ramus, 2.3 times as long as proximal seta wich inserted closer to ventral side, and about 1.3 times as long distal lateral seta. Inner apical seta smooth and slender, inserted closer to ventral surface, about 0.35 times as long as ramus. Middle apical seta strongest, inserted distally, without breaking plane, unipinnate, with slightly curled tip, about 1.6 times as long as outer apical seta and 0.2 times as long as whole body. Outer apical seta also strong, without breaking plane, unipinnate distally, but inserted closer to ventral side than middle apical seta, about 0.75 times as long as ramus.
Antennula ( Fig. 7D View FIGURE 7 ), antenna ( Fig. 7A View FIGURE 7 ), mouth appendages ( Fig. 7E View FIGURE 7 ), and first two pairs of swimming legs ( Fig. 7A View FIGURE 7 ) as in K. lakewayi .
Third swimming leg ( Fig. 8B View FIGURE 8 ) also very similar to that in K. lakewayi but apophysis nearly flat, with narrow notch on outer side and shallow notch on distal margin, making distal part into blade-like structure; ornamentation same as in K. lakewayi , but longitudinal row of large spinules on outer margin missing; exopodal spine also 1.5 times as long as apophysis, but with less pronounced inner hump in proximal part.
Fourth swimming leg very similar to next two species (see below), with five small spinules on basis at base of endopod (but not as close to endopod as in K. lakewayi ), and endopod with seven (on right leg) or eight (on left leg) large spinules arranged into scoop-like structure. Apical seta on third exopodal segment 1.3 times as long as entire exopod and three times as long as outer spine.
Fifth leg ( Fig. 7C View FIGURE 7 ) without any difference from that in K. lakewayi , except for several shallow cuticular pits on anterior surface in proximal part, and very weak outline of cuticular window in some specimens.
Sixth legs ( Fig. 7C View FIGURE 7 ) as in K. lakewayi .
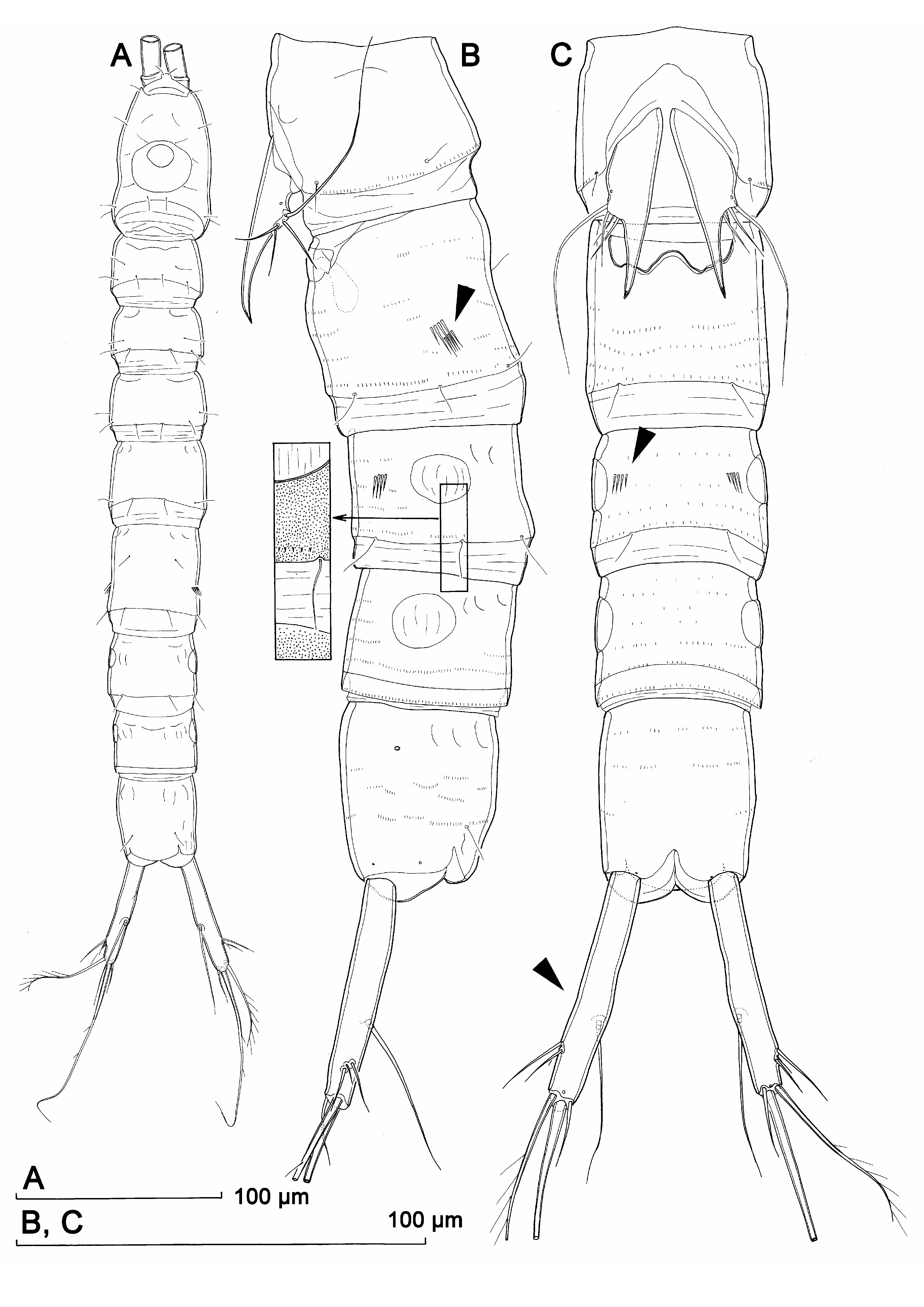
Female (based on allotype and several paratypes). Body length from 383 to 438 µm (402 µm in allotype). Habitus ( Fig. 9A View FIGURE 9 ), ornamentation of prosomites, colour, and nauplius eye similar to those in male, except genital and first abdominal somite fused into double-somite and middle part slightly less slender. Prosome/urosome ratio 0.7; greatest width from dorsal view hard to establish; body length/width ratio 8.8; cephalothorax only slightly wider than genital double-somite.
Genital double-somite ( Figs. 8D View FIGURE 8 , 9B, C View FIGURE 9 ) about 1.2 times as long as wide in dorsal view, without any trace of subdivision except for pair of ancestral dorso-lateral sensilla at middle; additionally ornamented with six posterior sensilla (two dorsal, two ventral and two lateral), numerous transverse rows of minute spinules, and two parallel short rows of four large spinules in posterior half laterally (arrowed in Fig. 9B View FIGURE 9 ). Genital complex ( Figs. 8D View FIGURE 8 , 9B, C View FIGURE 9 ) as in K. lakewayi , except outer distal corners of genital operculum not produced into spinifom processes.
Third urosomite ( Fig. 9B, C View FIGURE 9 ) similar to that in male, with two groups of four large spinules ventrally (arrowed in Fig. 9C View FIGURE 9 ); lateral cuticular windows well developed and highly visible.
Fourth (preanal), and fifth (anal) urosomites also very similar to those in male ( Fig. 9B, C View FIGURE 9 ), without any large spinules except those inside anal sinus.
Caudal rami ( Figs. 8C View FIGURE 8 , 9A, B, C View FIGURE 9 ) similar to those in male, but slightly inflated at middle in ventral view (arrowed in Fig. 9C View FIGURE 9 ) and more divergent.
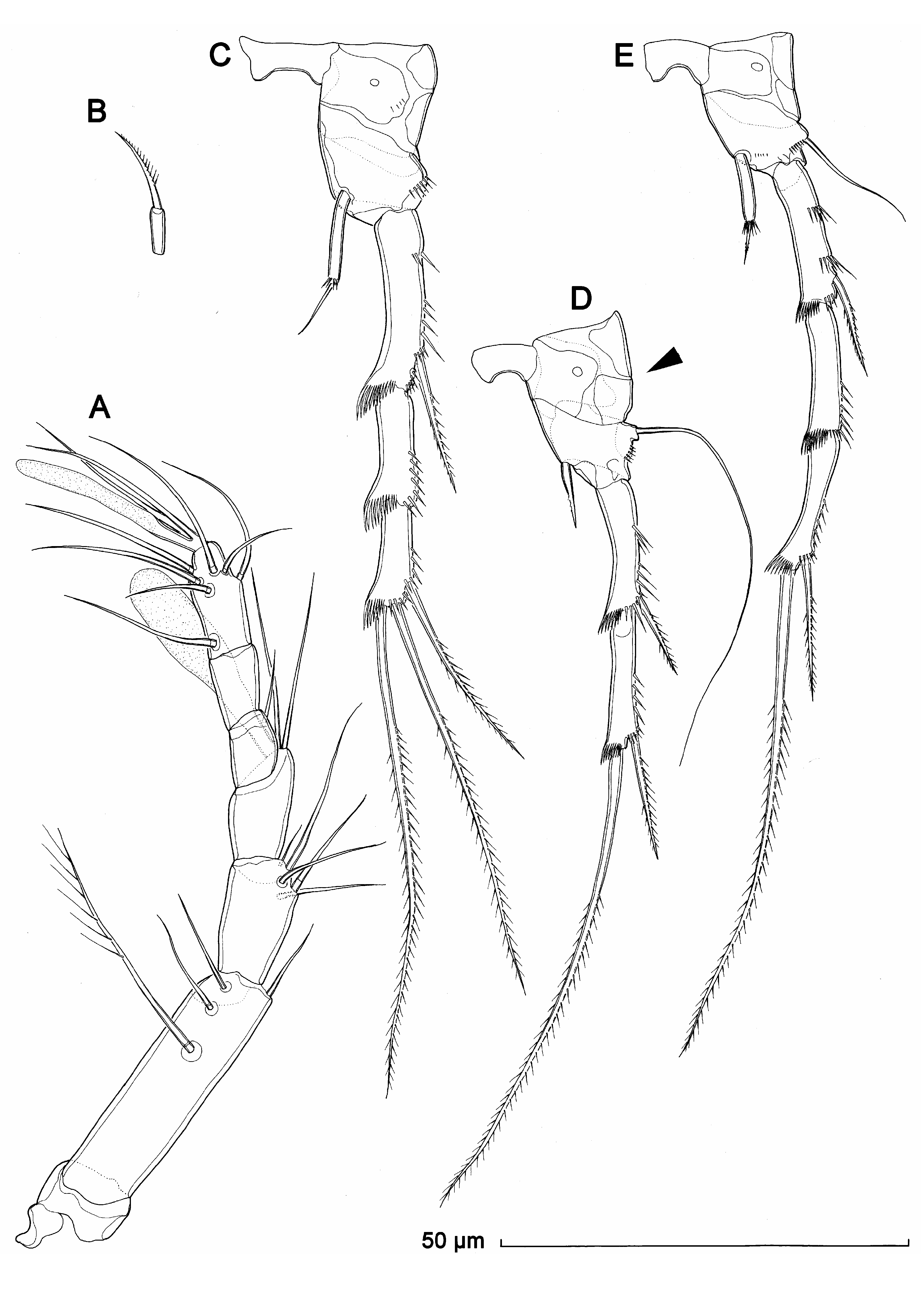
Antennula ( Fig. 10A View FIGURE 10 ) very similar to that in K. lakewayi , with slightly more robust aesthetascs.
Antenna ( Fig. 10B View FIGURE 10 ), mouth appendages ( Fig. 8E View FIGURE 8 ), first swimming leg ( Fig. 8E View FIGURE 8 ), second swimming leg ( Fig. 10C View FIGURE 10 ), and exopod of fourth swimming leg ( Fig. 10E View FIGURE 10 ) very similar to those in male and without any difference from those in K. lakewayi .
Endopod of second swimming leg ( Fig. 10C View FIGURE 10 ) 6.7 times as long as wide, its apical seta 0.7 times as long as segment.
Third swimming leg ( Fig. 10D View FIGURE 10 ) similar to K. lakewayi , but with more strongly developed cuticular plates on praecoxa and coxa (arrowed in Fig. 10D View FIGURE 10 ), and with proportionately longer apical seta on second segment and shorter endopod.
Endopod of fourth swimming leg ( Fig. 10E View FIGURE 10 ) about five times as long as wide, half as long as first exopodal segment, armed with single slender bipinnate seta apically; ornamented with six slender spinules along distal margin, at base of apical seta. Exopod similar to that of male.
Fifth leg ( Figs. 8D View FIGURE 8 , 9B View FIGURE 9 ) very similar to that of male, only slightly more elongated, with narrower distal part and cuticular window on anterior surface better defined, but not observable under light microscope.
Sixth leg ( Figs. 8D View FIGURE 8 , 9C View FIGURE 9 ) vestigial, both fused into simple bilobate cuticular plate, covering gonopore, unornamented and unarmed; outer distal corners not produced into sharp processes, but well rounded and shorter than inner lobes.
Etymology. The species name comes from its type locality (bore SB spelled in English), but should be treated as comprising an arbitrary combination of letters that can be treated as a Latin word and may be conceived as a noun in apposition to the generic name.
Variability. Body length in males ranges from 374 to 425 µm (395 µm average; n = 4), and from 383 to 438 µm in females (405 µm average; n = 15). Note that males are slightly smaller than females, but that may be a consequence of a low number (four) of the male specimens collected and examined. The endopods of the male fourth leg have always seven spinules on the right leg and nine on the left one, which is an unusual asymmetry, but it seems to be a character present also in the next two species (described below). The shape of the apophysis of the third leg in male is not variable, and it is a very good morphological character, as are the groups of large spinules on the urosome of females. The very low morphological variability recorded in this species is not surprising, considering that the species was collected from a single bore, which was the only access at the time of sampling to a small calcrete body in south-east of the Yeelirrie area (see Fig. 24 View FIGURE 24 ).
Remarks. Kinnecaris esbe sp. nov. has the longest and most slender caudal rami of all the Australian congeners, which are also among the most slender recorded for the family. In every other aspect, this is a typical member of the genus Kinnecaris Jakobi, 1972 . Morphology would suggest that this species is most closely related to two other short range endemics from Yeelirrie: K. linel sp. nov. and K. uranusi sp. nov. (see below). Unfortunately, despite repeated attempts, we were not able to get any COI sequences of K. esbe to test this relationship by molecular methods. Besides shorter caudal rami, the latter two species differ from K. esbe in the ornamentation of the genital double-somite in female (only one row of large spinules laterally; arrowed in Fig. 11B View FIGURE 11 ), and to a smaller extent in the shape of the third leg apophysis in male. Relying only on morphological evidence, we would speculate that these three species represent a monophyletic group among Australian Kinnecaris members (and also on a global scale), but molecular results disprove a close relationship of K. linel and K. uranusi (see discussion further below).
| WAM |
Western Australian Museum |
| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.