
Issiodoromys pauffiensis Vianey-Liaud, 1976
|
publication ID |
https://doi.org/ 10.5252/g2014n4a4 |
|
DOI |
https://doi.org/10.5281/zenodo.4837352 |
|
persistent identifier |
https://treatment.plazi.org/id/03D6987B-447E-B139-FF04-1440FF5BE088 |
|
treatment provided by |
Felipe |
|
scientific name |
Issiodoromys pauffiensis Vianey-Liaud, 1976 |
| status |
|
Issiodoromys pauffiensis Vianey-Liaud, 1976
HOLOTYPE. — Left dentary with p4-m3, MPF-1 (Vianey- Liaud 1976)); coll. UM2 .
TYPE LOCALITY. — Mas-de-Pauffié, Lot, Quercy, MP 26 (Chattian).
EMENDED DIAGNOSIS. — (Completing Vianey-Liaud 1976: 55 and Schmidt-Kittler & Vianey-Liaud 1987): Evolutionary stage intermediate between I. minor (Schlosser, 1884) and I. quercyi (Schlosser, 1884) . Cement rarely present at the bottom of sinus and sinusid. Horizontal uppermasseteric crest strongerthan in I. minor , especially in its posterior part.
Differs from I. minor by the more elongated anterior lobe of dp4, including two well-developed oblique ridges, mesially to the mesolophid (Ldp4 of I. pauffiensis from Mas-de-Pauffié = 2.71 mm to 3.01 mm); antesinusid generally absent on molars, anteroconid reduced to a weak vertical ridge when present, and in some specimens preceded by a small additional inflection of the crown on dp4; enamel cover thinner; simplified occlusal patterns in some individuals. Differs from I. quercyi by itssmaller size and relatively frequent remnants of the antesinusid-anteroconid on molars.
Differs from I. limognensis Schmidt-Kittler & Vianey- Liaud, 1987, in having the anterior lobe of dp4 less stretched, including less pronounced antesinusids (Ldp4 of I. limognensis from Pech-Desse = 2.96 mm to 3.67 mm), cement limited to the bottom of sinussinusid when present, whereas it reaches the occlusal surface – including teeth slightly to moderately worn – in I. limognensis .
MATERIAL EXAMINED. — 29 DP4 ( SPV 200-228), 40 P4 ( SPV 229-266), 113 M1-2 ( SPV 267-389), 26 M3 ( SPV 390-414); 29 dp4 ( SPV 415-442), 21 p4 ( SPV 443- 463), 101 m 1-2 ( SPV 464-564), 23 m 3 SPV 565-589).
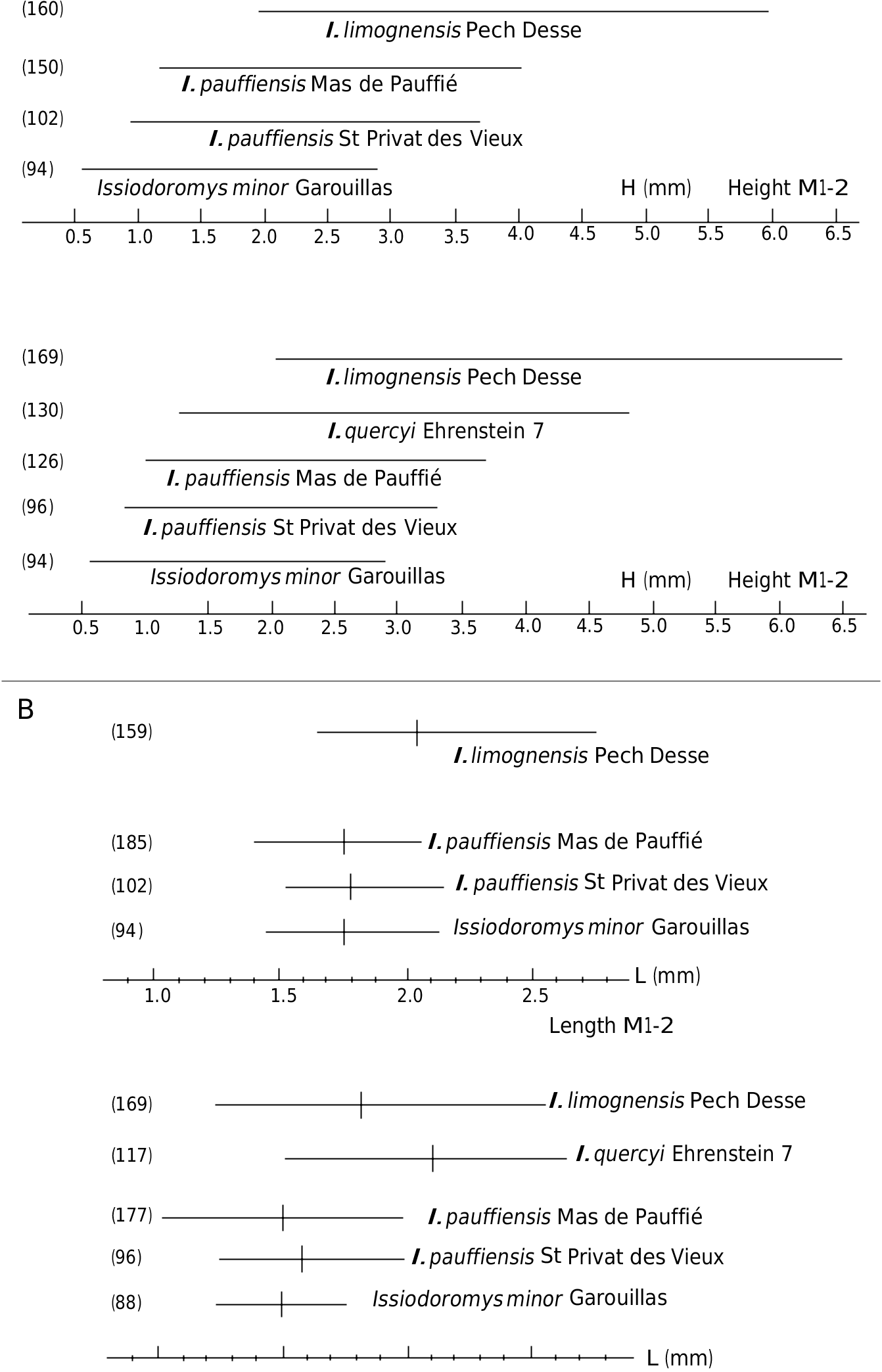
MEASUREMENTS. — To the variations in size between the first and second molars (the first being generally smaller than the second) must be added those due to effects of wear on the occlusal surface orientation of hypsodont teeth (Vianey-Liaud et al. 1995: 274, fig. 12), and finally those due to individual variability. Ŋe size variations are expressed by the Pearson coefficient: v = 100Sd / Lmoy ( Table 2 View TABLE ). Ŋe low size variability observed at Saint-Privat-des-Vieux reinforces the hypothesis on the taphonomy of the fossil assemblage from this locality ( Bonnet et al. 2011: 12).
DESCRIPTION
Lower teeth
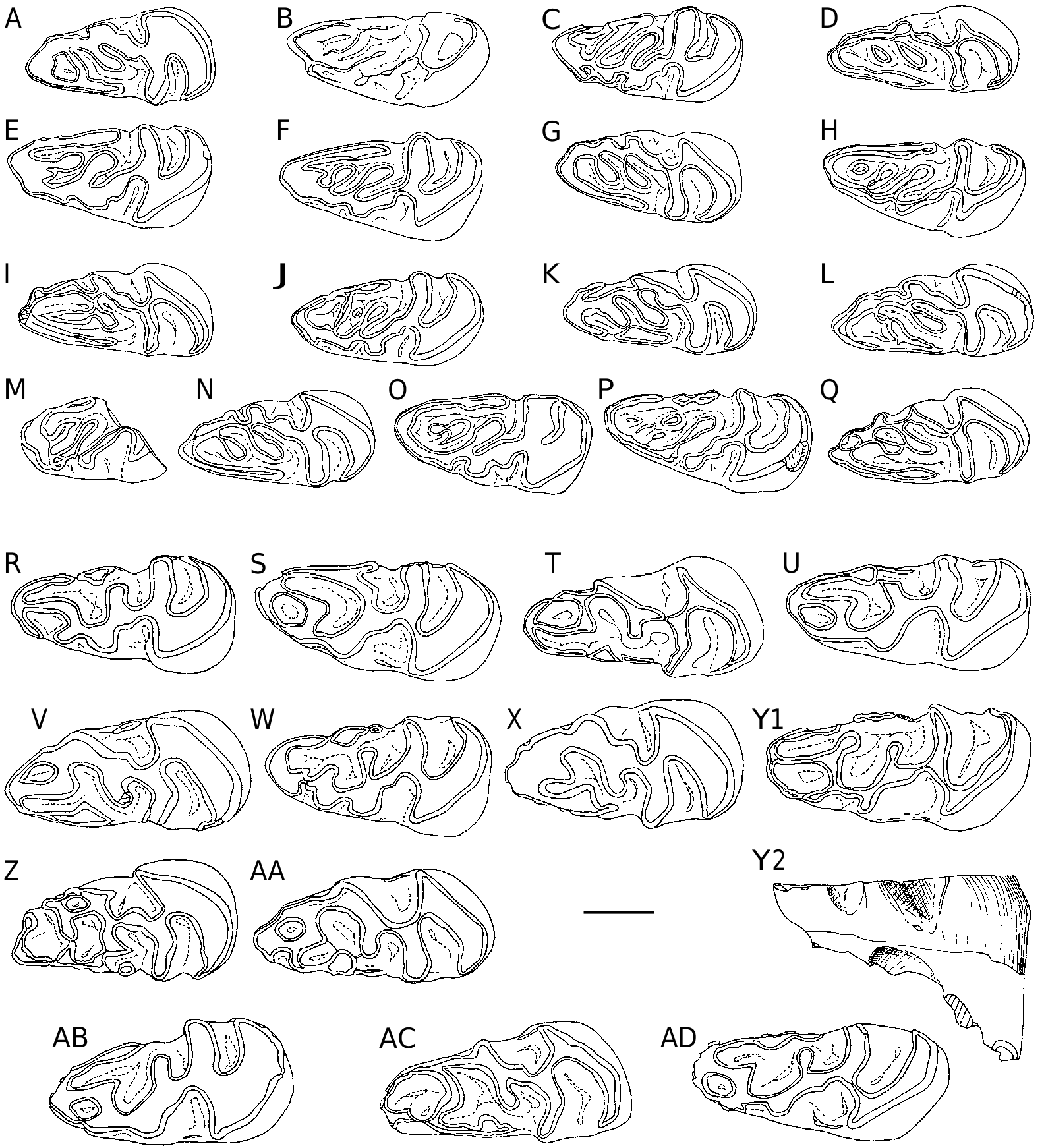
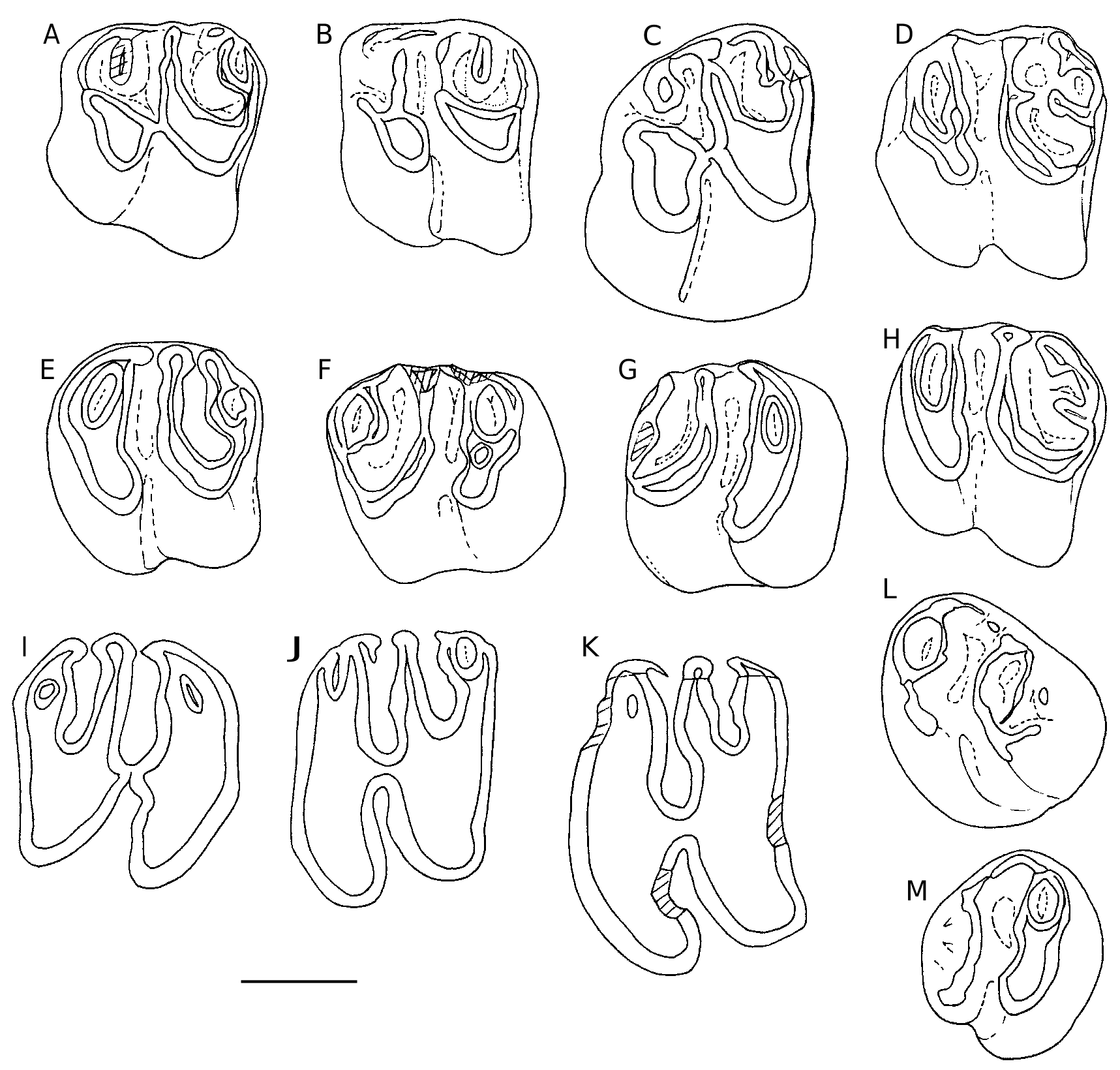
dp4 ( Fig. 4 View FIG ). Ŋe morphology of dp4 posterior lobe is constant, while some variations are observed in the anterior lobe. Ŋis is actually the part of dp4 that most changes during the evolution of the lineage, extending mesially the abrading surface of the tooth row. All dp4 show 2-3 oblique ridges between the mesolophid and the prelobe tip. Ŋe protoconid is split by a small labial notch, already sketched in populations of I. minor and present in the type population of I. pauffiensis (e.g., Schmidt-Kittler et al. 1997: 385, fig. 7; Vianey-Liaud et al. 1995: 267, fig. 5). Ŋe elongated central cuspid, which occupies the mesolophid position, runs obliquely towards the anterior arm of the protoconid to which it may be connected or not. Once upon 20 well preserved teeth, this central cusp reaches the tip of the prelobe. In all other cases, it is preceded by two oblique crestids parallel to it. Ŋe first is usually reduced to a small cusp. A continuous cingulid, as high as other ridges, follows the outline of the anterior lobe of teeth from the front flank of the
1.31 0.138 1.42 protoconid to the antero-lingual side of synclinid III (SIII). Labially, this cingulid shows two constant inflections: the antesinusid ahead of the protoconid, and a generally shallow pre-antesinusid.Ŋe lingual portion of the cingulid is vertically flat, and starts in continuity with the anterior cuspid of prelobe. Ŋis cuspid is prominent on unworn teeth. Ŋe size and morphology of these dp4 are very similar to that of dp 4 type population ( Table 2 View TABLE ).
p4 ( Fig. 5 View FIG A-C). Ŋere is a low morphological variability. A short mesio-distal expansion starting from the anterolophid inward SII is rarely observed (4/20). A small cingulid may be present at the base of the lingual opening of SIII (5/20). A p4 (1/20) bears only a remnant of the anteroconid.
m1-2. Ŋe vestigial anteroconid / antesinusid – as a mesiolabial vertical bulge – seems more frequent on molars than on p4, but this may be due to the more important number of m1-2. On little worn m1-2, 18/77 teeth have this vestige, while it is absent (or lost due to abrasion) on 23 worn teeth. Fourteen teeth exhibit a short mesiodistal spur in SII. On a single specimen, a mesiodistal spur is added to that in the SIV. Another specimen shows a lingual interruption of the mesolophid, and displays two small lin- gual protrusions, the most distal one blocking the lingual opening of the SIII ( Fig. 5E View FIG ). Ŋe only very worn tooth (H = 0.77 mm) exhibits a thin layer of cement at the bottom of the sinusid ( Fig. 5I View FIG ); this cement is not visible on the other teeth.
m3 ( Fig. 5 View FIG J-L). Ŋe mesiodistal spur ( Fig. 5L View FIG ) is present in the SII of 5/14 teeth and the anteroconid is detectable on one tooth only.
Upper teeth
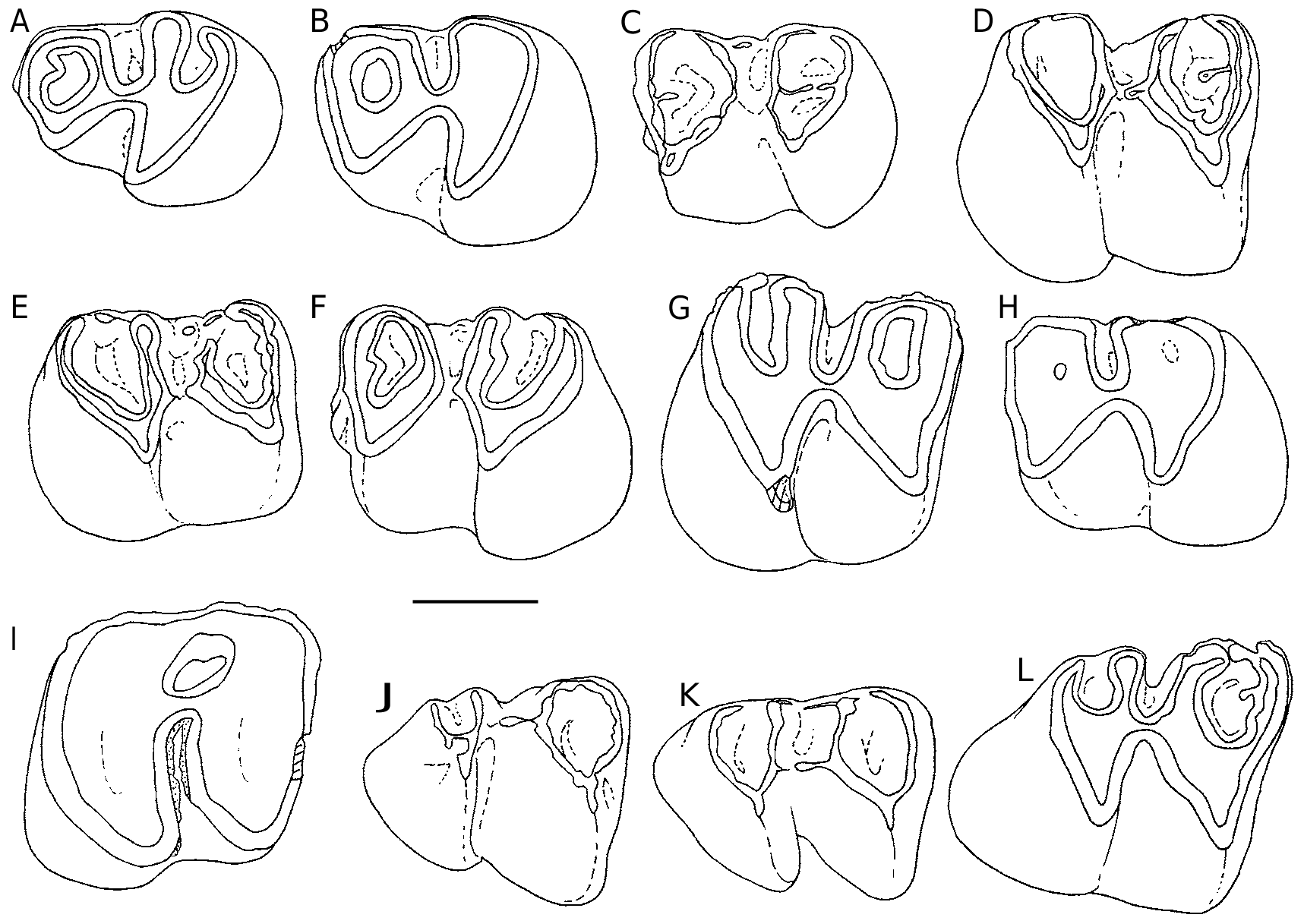
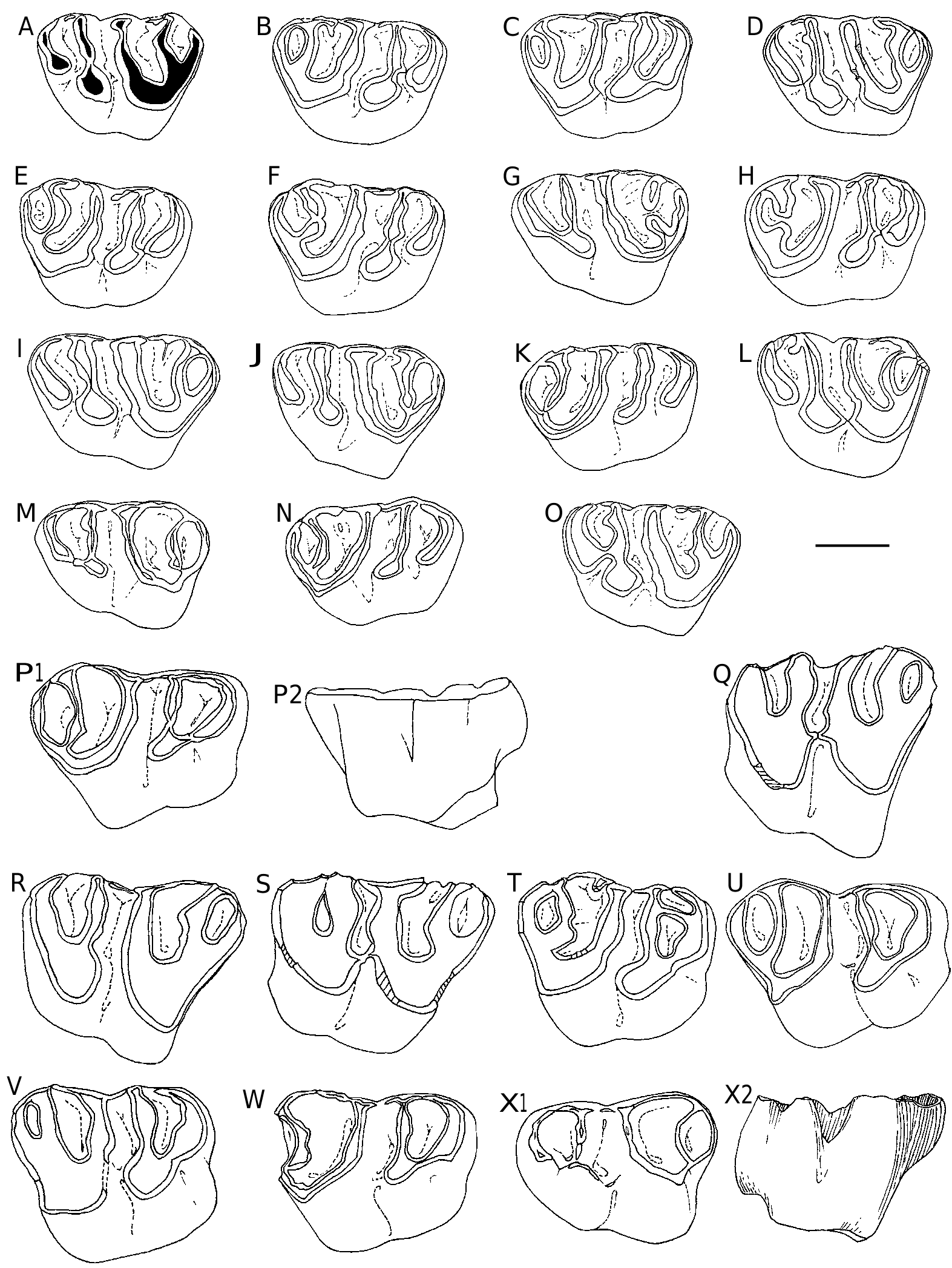
DP4( Fig. 6 View FIG ). Again morphological variations are minimal ( Fig. 6 View FIG A-O). Ŋe anticline 4 may be continuous (21/24) or split into two parts (3/24). We observe a light variability in the labial cingulum blocking the opening of the median syncline (SII) (e.g., Fig. 6C View FIG ). It is sometimes absent and sometimes high, resulting of proximity of two spurs coming from the labial ends of the anticlines 2 and 3, or only one labial spur from the anticline 2, mesiodistal or oblique. Ŋe buccal cingular edge of the syncline III is usually swollen as a cusp that can spawn a spur extending into the syncline. P4 ( Fig. 7 View FIG A-C). Ŋe lingual end of the anticline 4 is free (12/29) or connected to the posterior cingulum (17/29). Ŋe labial portion of the anterior cingulum is rarely complete (1/29), it is either vestigial (11/29) or absent. Ŋe paracone is sometimes isolated from the protocone (7/29) due to the interruption of the protoloph (anticline 2).
M1-2 ( Fig. 7 View FIG D-K). the morphological variations are minor and mainly concern SIV, which is mostly closed (98/101), or rarely mesially opened (2/101). A small mesiodistal spur runs backwards from the anticline 3 in the SIII (1/101).
M3 ( Fig. 7L, M View FIG ). Ŋe size and shape of the M3 are more variable than those of M1-2, especially
because of the greater or lesser reduction in the posterior lobe. On one of the smaller specimens, it only remains the anticline 3 at the back of the tooth.
DISCUSSION
Since the description of I. pauffiensis (Vianey-Liaud 1976) , we collected additional material for its type locality; for the present work, it has been measured, observed and compared to that of Saint-Privat-des-Vieux. Molars from Saint- Privat-des-Vieux are similar in size to those of I. pauffiensis from Mas-de-Pauffié. Ŋeir size is significantly smaller than that of I. quercyi (Ehrenstein 7, Germany [ Fig. 6 View FIG P-Z; Fig. 8B View FIG ], and Sarèle, France ; MP 27) ( Table 2 View TABLE ).
Ŋe size variability (length and width of teeth) of I. pauffiensis from Saint-Privat-des-Vieux is not higher than that observed in extant populations of brachyodont rodents or other mammals (e.g., Vianey-Liaud & Legendre 1986: 921, table 1). Ŋis variability is also less than that observed in fossil rodent teeth populations displaying a strong asymmetric hypsodonty, from both fissure fillings and stratified sedimentary deposits (e.g Vianey-Liaud & Legendre 1986: 921, table 3). It is thus a little lower than that of the I. pauffiensis type population from Mas-de-Pauffié, or that of I. quercyi from Ehrenstein 7 ( Table 2 View TABLE ; Fig. 8B View FIG ). Ŋe range of crown height is within that of species exhibiting similar to close hypsodonty (the maximum and minimum heights being slightly lower than those from Mas-de-Pauffié, and slightly higher than in I. minor from Le Garouillas). Ŋis range is clearly wider for I. quercyi from Ehrenstein 7, and also for I. limognensis from Pech Desse, both species being significantly more hypsodont ( Fig. 8A View FIG ).
Morphological variability observed in Mas-de- Pauffié, especially for deciduous teeth, is the same as described above for Saint-Privat-des-Vieux. Minor differences can be noted with regard to the dp4. Morphological variation in 25 dp4 from Mas-de-Pauffié shows the same characteristics as in the population of Saint-Privat-des-Vieux, except the presence of a low cingulid which partially or completely closes the bottom of the opening of the SIII on five specimens. Ŋis cingulid is also occasionally present on p4 from both localities or on molars: its absence on these five dp4 may be a statistical bias.
Postcranial elements of I. pauffiensis from Saint-Privat-des-Vieux – in particular astragali, calcanei and humeri fragments – are quite similar to those of Mas-de-Pauffié (Vianey-Liaud et al. in press).
| MP |
Mohonk Preserve, Inc. |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |