
Haliclona (Flagellia) porosa ( Fristedt, 1887 ) Van Soest, 2017
|
publication ID |
https://doi.org/ 10.5852/ejt.2017.351 |
|
persistent identifier |
https://treatment.plazi.org/id/510FEC4E-8B53-904E-FE63-FA401064F80A |
|
treatment provided by |
Carolina |
|
scientific name |
Haliclona (Flagellia) porosa ( Fristedt, 1887 ) |
| status |
subgen. et comb. nov. |
Haliclona (Flagellia) porosa ( Fristedt, 1887) View in CoL subgen. et comb. nov.
Desmacella porosa Fristedt, 1887: 440 , pl. 24 figs 36–37, pl. 28 fig. 15.
Gellius vagabundus Vosmaer, 1885: 29 View in CoL , only the var. γ, pl. V figs 36–38.
? Gellius rhaphidiophorus Brøndsted, 1933: 18 View in CoL , fig. 7.
Gellius porosus View in CoL – Lundbeck 1902: 73, pl. XIV fig. 2. — Lundbeck 1909: 434. — Hentschel 1916: 11. —
Ferrer Hernandez 1918: 22, fig. 3. — Ferrer Hernandez 1923: 16. — Rezvoi 1924: 243. — Rezvoi
1928: 91. — Hentschel 1929: 978. — Koltun 1959: 213. Haliclona porosus View in CoL – Koltun 1962: 186. — Hoshino 1987: 38. Gellius flagellifer View in CoL – Lambe, 1896: 185, pl. I fig. 4. — Topsent 1896: 281, pl. VIII fig. 4. Hemigellius View in CoL sp. aff. flagellifer View in CoL – De Weerdt & Van Soest 1987: 315.
non Gellius flagellifer sensu Koltun 1959: 212 View in CoL (= Haliclona (Gellius) View in CoL sp.); nec: Hemigellius View in CoL sp. aff. flagellifer sensu Ginn et al. 1998: 1099 View in CoL (= Haliclona (Gellius) View in CoL sp.)
Material examined
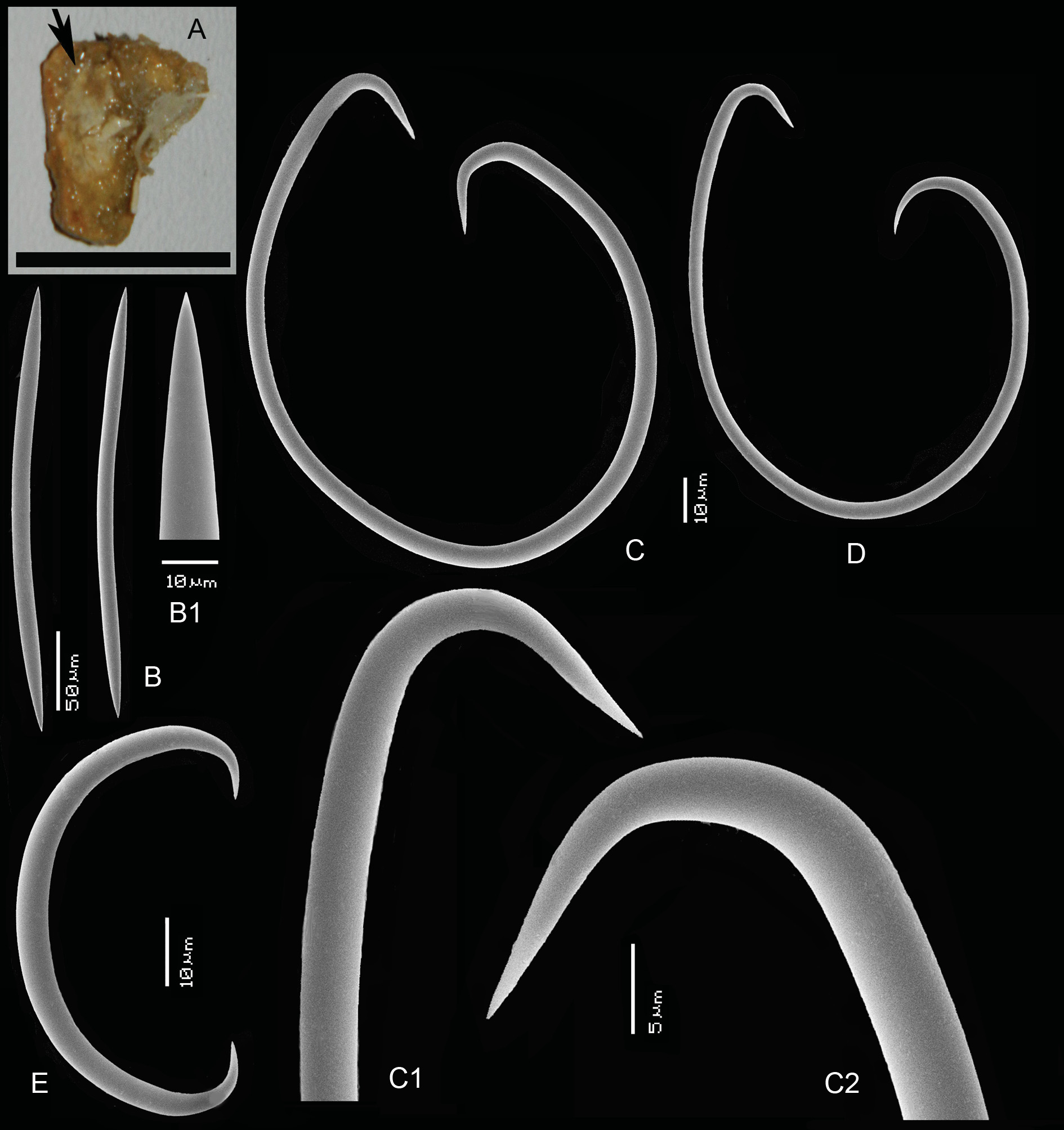
BARENTS SEA: slide only ( Fig. 12 View Fig A–F), Willem Barents Expedition,?1880, depth uncertain, between 212 and 297 m ( ZMA Por. 20742).
MAURITANIA: off Banc d’Arguin, 114 m depth, Van Veen grab, coll. R.W.M. Van Soest, Mauritania II Exped. stat. 033, 9 Aug. 1988 ( ZMA Por. 06624) ( Fig. 13 View Fig A–F).
Description
Because of the substantial difference between the two localities, each specimen is described separately.
ZMA 20742 ( Fig. 12 View Fig A–F) is a slide only, made from a dried fragment apparently no longer present in the collection.
ZMA 06624 ( Fig. 13A View Fig ) is a small encrustation of 5 × 5 × 2 mm on dead Lophelia corals, surface optically smooth. Consistency soft.
SKELETON. Specimen 20742 ( Fig. 12A View Fig ): the surface has confused tangentially arranged single spicules.The choanosomal skeleton has paucispicular ascending tracts interconnected irregularly by single spicules. Microscleres, especially normal sigmas, are relatively rare throughout the skeleton. Specimen 06642: no recognizable surface skeleton. The choanosomal skeleton is irregular, anisotropic with paucispicular
ascending tracts and single connecting spicules. Microscleres, especially the normal sigmas, are rare throughout the choanosome.
OXEAS. Straight or slightly curved, elongately cigar-shaped. ZMA 20742 ( Fig. 12A View Fig ): 243– 271 –297 × 8– 9. 7 –12 μm. ZMA 06624 ( Fig. 13B, B View Fig 1 View Fig ): 267– 307 -333 × 8.5– 11. 2 –13 μm.
2
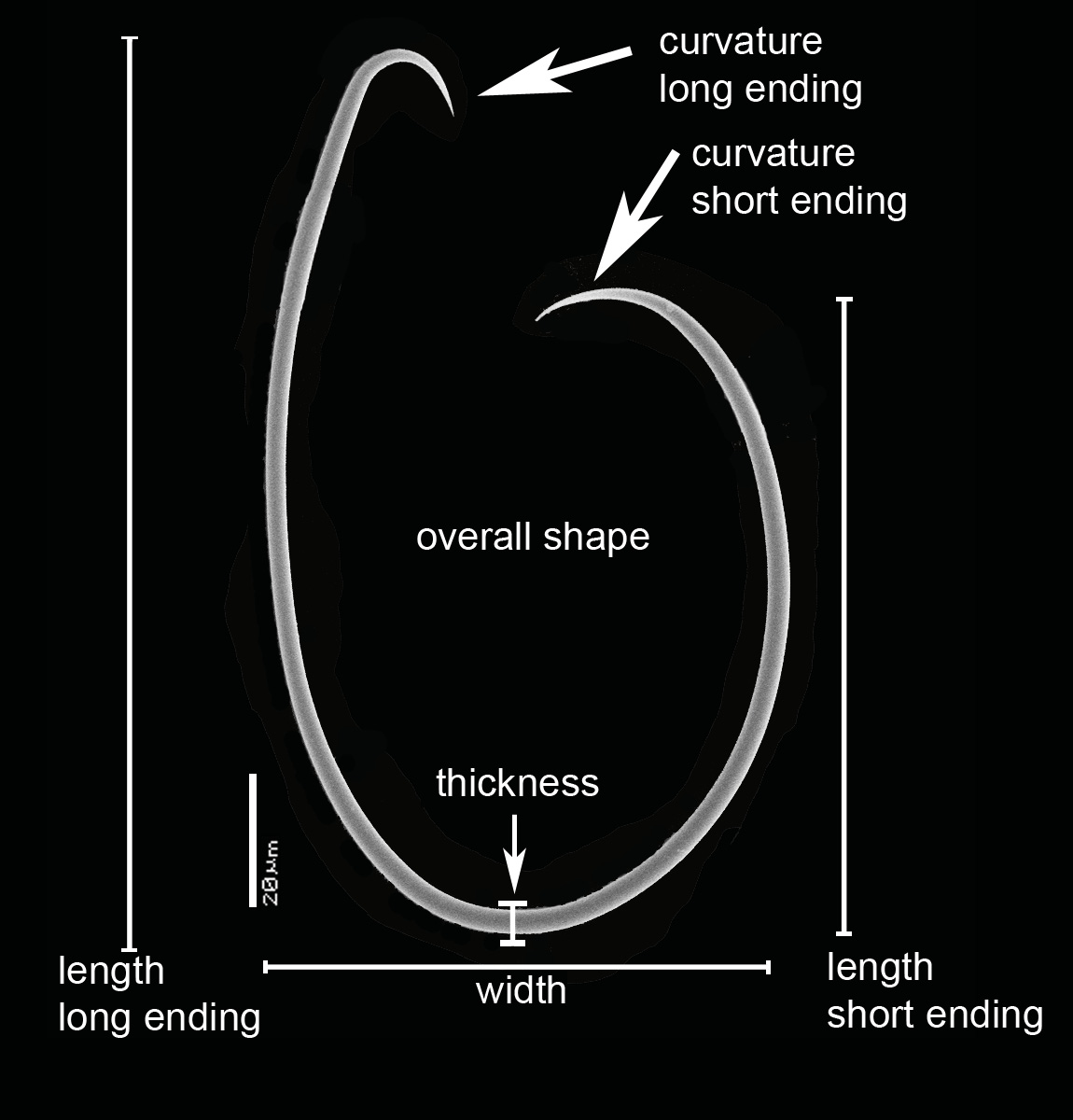
FLAGELLOSIGMAS. ZMA 20742 ( Fig. 12 View Fig B–F): elliptical to ovoid, in a large size range, but not divisible into two categories, length of long ending 48– 82 –108 μm, of short ending 27– 47 –59 μm, width 32– 46 –57 μm, thickness 1– 1. 8 –3 μm. ZMA 06624 ( Fig. 13C, C View Fig 1 View Fig , C 2 View Fig , D): predominantly ovoid or circular, larger than those of 20742, also in a large size range, not divisible. Long endings have straight curvature, length of long endings 57– 109 –156 μm, short endings 43–79–106 μm, width 42– 83 –117 μm, thickness 1.5– 3. 3 –5 μm.
NORMAL SIGMAS. Rare, in both. ZMA 20742: 45–48 × 2–2.5 μm (n=3). ZMA 06624 ( Fig. 13E View Fig ): 40– 51 – 61 × 2– 4. 0 –5 μm (n=9).
Distribution and ecology
Arctic waters, NW Pacific, Gulf of Biscay, Gulf of Saint Lawrence, off Mauritania (Marine Ecoregions West Greenland Shelf, East Greenland Shelf, North and East Barents Sea, South European Atlantic Shelf, Gulf of Saint Lawrence, Sahelian Upwelling). Depth occurrence 90– 400 m.
Remarks
All slides and specimens in the ZMA and RMNH collections labeled as Gellius vagabundus remaining from the material studied by Vosmaer (1885), including several labeled as the var. γ, and even one specimen (nr. 74) indicated by Vosmaer (1885: 29, pl. V figs 38–38) as having the spiculation depicted, were examined. No flagellosigmas were found, all specimens and fragments belonged either to Desmacella , Hymeniacidon or Hemigellius , with sigmas of normal shape, or lacking. There is one dried sample without identification in the ZMA collection, bearing only a small label with text ‘Sp. XXX No. 76’, the number given to Gellius vagabundus by Vosmaer (1885). The sample consists of three fragments, all of which are Hymeniacidon -like (with larger and smaller styles, as depicted in pl. V figs 32–33). However, the ZMA 20742 slide presumably made from the dried material does have the spicules depicted in Vosmaer’s Pl. V figs 36–38. Although the number is 76, not 74, it is clear that this slide was made from a previously present dried fragment and is now is all that remains of Vosmaer’s var. γ.
Both the Barents Sea slide and the Mauritanian specimen have been assigned to H. (F.) porosa based on spicule shapes, and the presence of a single category of normal sigmas that are characteristically rare. They resemble Fristedt’s description of Desmacella porosa from Davis Strait, although the type specimen – listed to be present as Gellius porosus in the Zoologisk Museum Copenhagen under reg. nr. DEM 107 – was much larger (9 × 6 cm). The size of the oxeas was given by Fristedt as 350 μm, but Lundbeck (1902) re-examined the type and found some of them to be as small as 250 μm. Fristedt did not mention any normal sigmas, but Lundbeck (1902) found several normal sigmas. The length of the flagellosigmas was given by Fristedt as 120 μm across, somewhat larger than the ones of ZMA 20742, but smaller than those of ZMA 06624. Lundbeck (1902), in his description of a specimen from the N coast of Iceland, found oxeas and flagellosigmas in the same size range as those of the above described Barents Sea slide (ZMA Por. 20742), and the normal sigmas were 50–80 μm, somewhat larger than those of the present material.
Topsent (1896: 281, pl. VIII fig. 4) reported Gellius flagellifer from the nearby Gulf of Biscaye (depth 400 m), with oxeas 350 × 13–14 μm and flagellosigmas up to 90–100 × 2 μm. The drawing of the flagellosigmas closely resembles the present material. He neither discussed, nor figured the normal sigmas, suggesting that they were rare [he admitted to their presence later ( Topsent 1904)]. The rarity of the normal sigmas makes it likely that the material belongs to H. (F.) porosa . Lambe’s (1896) record of Gellius flagellifer is probably also H. (F.) porosa for the same reasons.
Ferrer Hernandez (1918: 22, fig. 3) reported Gellius porosus from the coast of Asturias, N Spain, at 200 m depth (material originally collected by Orueta). His description and figure may be similar to the type of Fristedt.
In addition to H. (F.) porosa, Lundbeck (as well as other authors, e.g., Rezvoi 1928: 91) also described Arctic specimens assigned to Gellius flagellifer Ridley & Dendy, 1886 . Lundbeck’s specimens differ from his H. (F.) porosa in having larger oxeas (up to 476 μm long), and abundant normal sigmas in a large size range. For these and other reasons, it is unlikely that these Arctic specimens are conspecific with specimens occurring in the Southern Ocean Prince Edward Archipelago. There are subtle small differences in the sizes and shapes of the flagellosigmas and the normal sigmas when compared with the type of H. (F.) flagellifera (see above). As Lundbeck insisted that these specimens were not conspecific with H. (F.) porosa (several authors, e.g., Topsent 1896 and Lambe 1896 thought otherwise), and access to the specimens was not possible, they remain provisionally as Haliclona (F.) spec. until their status as a separate species from H. (F.) porosa can be established (but see also below).
Brøndsted’s (1933) species Gellius rhaphidiophorus from Greenland at 410 m depth, was described as close to Gellius porosus , with ‟more or less flagelliform” sigmas of 71–110 μm, in addition to rather rare sigmas of 20–36 μm and raphides of 36–40 μm. Species with flagellosigmas and raphides are otherwise not known, so possibly the raphides are foreign, in which case it could be a junior synonym of H. (F.) porosa . However, conspecificity is uncertain because the presence of true flagellosigmas cannot be verified due to the lack of illustrations and the ambiguous description. Likewise, many records of Gellius porosus and Sigmadocia porosa (cf. above in the historical overview) remain to be substantiated, as they were either not taxonomically described or insufficiently characterized.
As mentioned above, Koltun (1959: 213) and Ginn et al. (1998: 1099) erroneously reported Gellius or Hemigellius flagellifer from Arctic and East Canadian waters, based on misidentification of their specimens.
The name combination Haliclona (F.) porosa is threatened by previous use of the name ‘ porosa ’ in combination with the unaccepted genus name Arcesios and the subgenus name Reniera . Arcesios porosa Duchassaing & Michelotti, 1864 is described unrecognizably and no original material is known to be extant in collections (cf. Van Soest et al. 1983). Schmidt (1870: 40) assigned this species to Reniera Schmidt, 1862 , claiming in a two-line comment that Arcesios was a junior synonym of Reniera , but failed to provide any evidence. Schmidt stated he had a specimen from ‟Crabb Island” [sic] that answered to the descriptions of A. porosa . He gave no description of this material and no specimen from ‟Crabb Island” is kept in the collections of the Museum of Comparative Zoology, Harvard, or the Musée de Zoologie at Strasbourg, where most of Schmidt’s 1870 material is housed. Although Reniera is a subgenus of Haliclona , there is no evidence that Arcesios porosa is a member of the subgenus. This fact precludes a definite and formal decision about the preoccupied state of the combination Haliclona (Flagellia) porosa and does not warrant proposal of a new name for it as a junior secondary homonym.
An additional homonym of the present combination is Reniera cinerea var. porosa Topsent, 1901 . This was renamed Reniera topsenti Thiele, 1905 because of Schmidt’s (1870) combination Reniera porosa (and was subsequently assigned to the genus Haliclona as H. topsenti by Burton (1940: 99). Because Schmidt’s combination has priority, even if it is likely not the same species as Arcesios porosa , the name Reniera topsenti and the current combination Haliclona topsenti remain accepted.


| ZMA |
Universiteit van Amsterdam, Zoologisch Museum |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Heteroscleromorpha |
|
Order |
|
|
Family |
|
|
Genus |
|
|
SubGenus |
Flagellia |
Haliclona (Flagellia) porosa ( Fristedt, 1887 )
| Van Soest, Rob W. M. 2017 |
Gellius flagellifer sensu
| Ginn B. K. & Logan A. & Thomas M. L. H. & Van Soest R. W. M. 1998: 1099 |
| Koltun V. M. 1959: 212 |
Gellius rhaphidiophorus Brøndsted, 1933: 18
| Brondsted H. V. 1933: 18 |
Gellius porosus
| Hentschel E. 1916: 11 |
| Lundbeck W. 1909: 434 |
| Lundbeck W. 1902: 73 |
Desmacella porosa
| Fristedt K. 1887: 440 |
Gellius vagabundus
| Vosmaer G. C. J. 1885: 29 |
Haliclona porosus
| Koltun 1962: 186 |
| Hoshino 1987: 38 |
| Lambe, 1896: 185 |
| Topsent 1896: 281 |
| De Weerdt & Van Soest 1987: 315 |